Agricultural Research Pesticides and Biofertilizers
OPEN ACCESS | Volume 6 - Issue 1 - 2026
ISSN No: 2994-0109 | Journal DOI: 10.61148/2994-0109/ARPB


Ehsanul Haque1 , Most Shanaj Parvin2, 3*
1North Dakota State University, Fargo, USA
2Leibniz University Hannover, Germany
3Bangladesh Agricultural Research Institute, Joydebpur, Gazipur, Bangladesh
*Corresponding Author: Most Shanaj Parvin, Bangladesh Agricultural Research Institute, Joydebpur, Gazipur-1701, Bangladesh.
Received: July 05, 2021
Accepted: July 10, 2021
Published: July 20, 2021
Citation: Md Ehsanul Haque and Most Shanaj Parvin. (2021) “Variations in Pathogenicity of three different forms of Rhizoctonia inoculum and Assessment of Cultivar Resistance of Sugar Beet.”, Journal of Agricultural Research Pesticides and Biofertilizers, 2(2); DOI:http;//doi.org/07.2021/1.1031.
Copyright: © 2021 Most Shanaj Parvin. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Rhizoctonia solani causes pre-emergence and post-emergence damping-off, as well as crown and root rot of sugar beet (Beta vulgaris L.), which significantly affects the yield returns in the USA and Europe. The pathogen overwinters as sclerotia or melanized mycelium. Traditionally, the resistance of cultivars to R. solani is evaluated by scoring disease reactions at the crowns and roots of older seedlings, thus resistance is not evaluated during seed germination. Moreover, earlier studies evaluated cultivars resistance to R. solani using colonized whole barley or wheat grains which, unlike sclerotia, are artificial inocula of the pathogen that require time, space and technical know-how to produce. Moreover, colonized grains are prone to contamination with other pathogens, consumed by rodents/birds while applied in the field, and are often uneconomic. Considering those limitations, a study was undertaken (1) to develop in vitro methods to generate large scale sclerotia, (2) to compare pathogenic potentials of sclerotia, mycelia, and colonized barley grains for optimization of damping-off assays, and (3) to evaluate Rhizoctonia resistance of selected commercial cultivars during the seed germination phase. Comparing six different culture media, we found that R. solani had the highest radial growth (8.9 ± 0.04, cm3) at 8-days and the maximum number of sclerotia produced (203 ± 4.6) at 28-days in CV8 medium. We demonstrated significant differences in pathogenicity of the three different forms of R. solani inocula and susceptibility of cultivars to pre- and post-emergence damping-off. The highest pre-emergence damping-off and root rot were observed with sclerotia, and the highest post-emergence damping-off was recorded with both sclerotial and colonized barley inocula. In addition, varietal differences in susceptibility to pre- and post-emergence damping-off were noted. The highest pre-emergence damping-off was recorded on cv Crystal 101RR and lowest in Maribo MA 504. The highest post-emergence damping-off was recorded on BTS 8500 and the lowest in Crystal 467. The maximum mean root rot was observed in BTS 8500, BTS 8606, and Crystal 101R. Our studies demonstrated that sclerotia serve as efficient natural inocula, reemphasized that host-pathogen interactions differ at the early vs. late stages of sugar beet growth, and highlighted the need to reevaluate commercial sugar beet cultivars for resistance at the seed germination stage.
Introduction:
Sugar beet (Beta vulgaris, L.) contributes approximately 20% of sugar worldwide (Dohm et al. 2014) (International Sugar Organization, 2018). The rest is mainly derived from sugarcane (Phillips et al. 2009). In the US, sugar beet contributes 55% of total sugar production (USDA, ERS, 2019). From sowing to post-harvesting, sugar beet is vulnerable to various biotic and abiotic stresses.
Many soil-borne pathogens, such as Rhizoctonia solani Kühn affect sugar beet stands and sugar yields. It is a genetically complex soil-borne fungus that causes pre-emergence and post-emergence damping-off, and also root and crown rot (Mukhopadhyay, 1987; (Windels and Nabben 1989); Harveson et al., 2009). There are 13 anastomosis groups (AGs) of R. solani, which consist of AG 1 to AG 13; some AGs are host-specific while others have wide host ranges (Carling et al. 2002; Parmeter et al. 1969). Sugar beet is prone to infection by the AG 2-2 strain (Parmeter et al. 1969). The main AG subgroup that seriously impacts sugar beet yield is AG 2-2 IIIB (Brantner and Windels 2009; Windels et al. 1997). However, the pathogenesis of this subgroup in sugar beet is not entirely well characterized. The primary inocula of the pathogen in nature are mycelia and sclerotia. This pathogen can survive in the soil for many years in the form of sclerotia which is an undifferentiated aggregation of thick-walled melanized cells (Sherwood 1967); (Adams and Papavizas 1970; Papavizas 1970). The sclerotia germinate under humid conditions and are often attracted by the root exudates of the germinated seedlings (Flentje et al. 1963). The host-pathogen interactions are generally initiated by mycelia that penetrate into the root cortex and cause infections to the tissue (Armentrout and Downer 1987; Christou 1962; Sneh et al. 1996). Rhizoctonia disease phenotyping in sugar beet fields often vary, primarily due to heterogeneous factors (biotic and abiotic), thus it is difficult to perform uniform disease trials (Bolton et al. 2010). To reduce variation in the experimental field, artificial inoculation using barley coated Rhizoctonia and greenhouse trials are usually performed as a complement to field trials. Preparing barley/wheat inocula requires time and space, technical know-how for large scale production, as well as protection from other air-borne pathogens. Moreover, it can be difficult to evaluate resistance levels of sugar beet cultivars outdoors that provide a good correlation between artificial inoculum and disease severity. Since R. solani rarely produces any basidiospores, it is difficult to use the same amount and concentration of fungal biomass each time (Cubeta and Vilgalys 1997). The utilization of sclerotia as inocula may circumvent those limitations. In an attempt to develop a uniform inoculation method and to better understand the R. solani-Beta vulgaris pathosystem, isolates of R. solani AG 2-2 IIIB were cultured in six different artificial media to study the morphology and development of sclerotia. Furthermore, infection severity was compared using three different forms of Rhizoctonia: sclerotia, mycelia, and colonized barley grains, in order to determine effective inoculation propagules and to evaluate the resistance response of the commercial cultivar in the greenhouse and in the field.
Materials and Methods:
Fungal Isolates of R. solani
Five R. solani isolates were collected from different diseased sugar beets from a field in Hickson, ND. Genomic DNAs (Norgen Biotek Corp, Fungi DNA Isolation Kit #26200) of the five isolates were used for polymerase chain reaction (PCR) with the internal transcribed spacer (ITS) (Sharon et al. 2008). Subsequently, PCR products were flushed by E.Z.N.A ®Cycle Pure Kit Omega Bio-tek, Norcross, GA) and four samples were sequenced by GenScript (Piscataway, NJ). The sequences were identical, and BLASTn analysis showed 100% sequence homology to R. solani AG 2-2 IIIB (Genbank accession: MN128569); thus, the five isolates were found to be clones of a single isolate. The isolate was maintained on CV8 and used for sub-culturing on six different media.
Culture Media and Development of Sclerotia of R. solani:
Microbial media play a significant role in the optimum mycelial growth and differentiation of different fungal species. The six different media: amended clarified V8 (CV8), 50% potato dextrose agar (PDA), 10% PDA, methylene- benomyl-vancomycin (MBV), cornmeal agar (CMA) and water agar (WA), were prepared following the “Manual of Microbiological Culture Media” (Difco & BBL Manual 2009). The experimental design was a complete block design with four replications. Mycelial discs (3 mm3) of R. solani, AG 2-2IIIB cut from the 7-day old mother colonies were transferred onto each of the six media in order to study the radial growth and development of sclerotia in vitro over time. Radial growth was measured using a digital caliper (Pittsburg 6” Composite DC, Item 93293) at four different time points: 2-day, 4-day, 6-day, and 8-day, respectively. Similarly, the number of sclerotia was counted at four-time points: 7-day, 14-day, 21-day, and 28-day, respectively. The experiment was repeated twice.
Categories of Different Sizes of Sclerotia:
Sclerotia were categorized into three different groups based on the size: large (≥4.00 mm), medium (≥2.00 mm), and small (≥0.5 mm). To evaluate the size effect of sclerotia on causing disease, an experiment was conducted in the humidity chamber at 35° C and 85% relative humidity. Three different categories of sclerotia were used to inoculate Crystal 101RR with a complete block design of four treatments (including non-inoculated check) and four replications. The total number of observations was 16 (4-treatment × 4-replication). Ten seeds were sown in each treatment. The seedling stand count was recorded at two weeks post-inoculation.
In vitro Inoculation on PDA Using Three Different Forms of R. solani inocula:
To compare the efficacies of the three different inocula (i.e., sclerotia, mycelia, and colonized barley seeds), sugar beet seeds were co-cultured with respective inoculum in PDA with four replications. A non-inoculated check with four replications was used as a control. Sugar beet seeds were washed with 70% ethanol for 1 minute and rinsed twice with sterile water. Seeds were dried on sterile blotter paper under the laminar airflow cabinet. Three seeds (seed to seed distance 1cm) were placed with sterile forceps on each culture plate and a different one of the three inocula was placed close to each seed on the plate. Germination data were recorded from 2 days post-inoculation (dpi) to 7 dpi. The experiment was repeated twice.
Greenhouse evaluation of cultivars’ susceptibility to Rhizoctonia inocula:
Trials were conducted in a greenhouse at North Dakota State University, Fargo, North Dakota, USA. The experimental layout was a Complete Block Design (CRD) with seven sugar beet cultivars, four treatments, and four replications. The total number of observations was 112 with a split-plot arrangement. The seven commercial cultivars screened in this investigation had the 2018 Rhizoctonia root rot ratings in parenthesis, as follows: Crystal 101RR (4.50), Crystal 467RR (3.94), Hilleshog 4302RR (3.71), Maribo MA 504 (4.25), BTS 8606 (4.24), BTS8500 (4.36) and BTS80RR52 (3.96) (sbreb.org/research/, Research Report 2018). Three different types of Rhizoctonia inocula: barley inoculum, sclerotia, and mycelia plug, as well as a non-inoculated check (autoclaved barley seed), were used for the inoculation of each cultivar. Plastic pots (27 x 13 x 13 cm3, T.O. Plastics, Inc.; Clearwater, MN, USA) were filled with vermiculite and perlite mixer (PRO-MIX FLX) amended with osmocote (15-9-12, 8˝ pot, 2.5 teaspoons per pot) fertilizer. Ten sugar beet seeds of each of the seven sugar beet cultivars were sowed into 2 cm deep furrow and 1 cm away from each seed in the center of each pot. Inoculation (one barley-inoculum/seed, one sclerotia/seed, one mycelial plug/seed, and mock inoculation) was done simultaneously with the sowed seed at the same depth and covered with mixer. The greenhouse temperature during the experiment was 25±2ºC during the day, with 80% relative humidity, and a 12-hour photoperiod. Plants were ensured water as needed to maintain adequate soil moisture conducive for plant growth and disease development.
Two weeks post-inoculation, the mean emergence and pre-emergence damping-off data were analyzed separately using split-plot R-package (Version 3.6.1, St. Louis, Missouri, USA). Percent post-emergence damping-off and stand counts data up to 42 dpi were also analyzed separately using split-plot R-package. At 56 dpi, plants were removed from pots, and roots were washed and rated for root rot disease severity using a modified 1-7 rating scale (Ruppel et al. 1979), where 1 = clean roots and no infection, 2 = < 5% of root surface with black/brown symptoms, 3 = 5-25% of root surface with black/brown symptoms; similarly, 4 = 26-50% , 5 = 51-75%, 6 = 75-100% of root surface with black/brown symptoms, and 7 = dead plants (withered). The data were subjected to analysis of variance (ANOVA) and Fisher’s Protected Least Significant Difference was used to separate root rot severity means using the split-plot R-package software (3.6.1) (Table 7).
Statistical analyses:
In both the in vitro and in vivo studies, the experiment was analyzed with the split-plot design using R-software (Version 3.6.1, St. Louis, Missouri, USA). Treatment means were distinguished by calculating Fisher’s Least significant difference at α = 0.05 confidence level.
Field evaluation of three different forms and rates of Rhizoctonia inocula:
Field trials were conducted at Hickson, North Dakota, USA. The experimental layout was a Randomized Complete Block Design (RCBD) using a susceptible cultivar, Crystal 101 RR, with ten treatments (sclerotial number 1, 2, 3; mycelial disc number 1, 2, 3; colonized barley number 1, 2, 3; and non-inoculated check) and four replications (i.e., total number of observations was 40). The row length and width were 30 feet by 11 feet, respectively. Three different forms and rates of Rhizoctonia inocula were applied immediately after the seeds were sown.
Results:
Effect of different culture media and days on the radial expansion:
Six different culture media were used to identify their effects on the radial growth of mycelia. There was a significant difference between the media, time points, interaction effects on the radial growth of mycelia, and the number of sclerotia. A mean comparison test was performed for radial growth and the number of sclerotia developed over different time points (Table 1 and Table 2).
|
Sources of variation |
Df |
Sum Sq |
Mean Sq |
F value |
Pr(>F) |
|
Block |
3 |
0.10 |
0.032 |
5.0218 |
0.01317 * |
|
Culture media |
5 |
33.45 |
6.690 |
1051.766 |
< 2e-16 *** |
|
Error a |
15 |
0.1 |
0.006 |
|
|
|
Time points/Days |
3 |
455.42 |
151.806 |
27906.532 |
< 2e-16 *** |
|
Culture media × Days |
15 |
6.10 |
0.406 |
74.7217 |
< 2e-16 *** |
|
Error b |
54 |
0.29 |
0.005 |
|
|
|
Significance. codes |
‘ ’Non-significant, ‘***’ 0.001, ‘**’ 0.01, ‘*’ 0.05 |
||||
|
cv(a) = 1.5 %, cv(b) = 1.4 %, Mean = 5.316667 |
|||||
Table 1: Analysis of variance for the effect of six culture media on radial growth (R. solani)
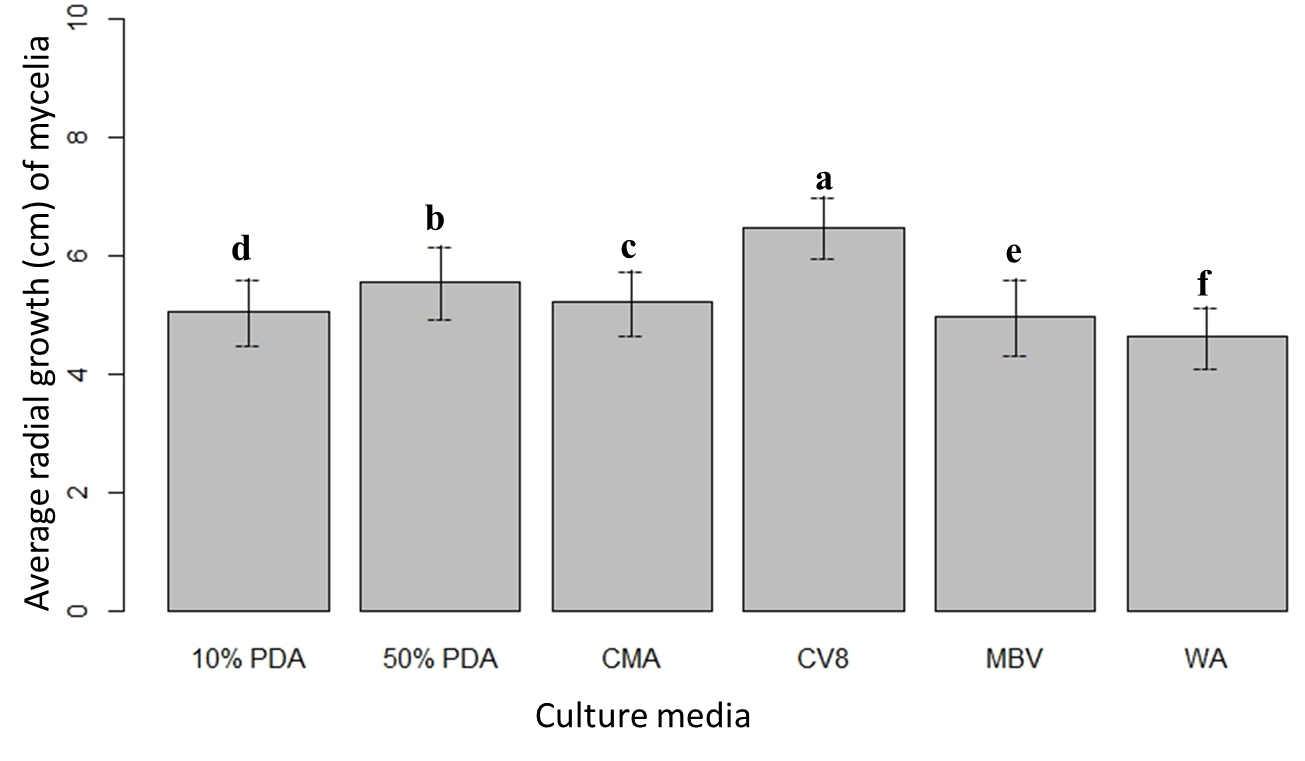
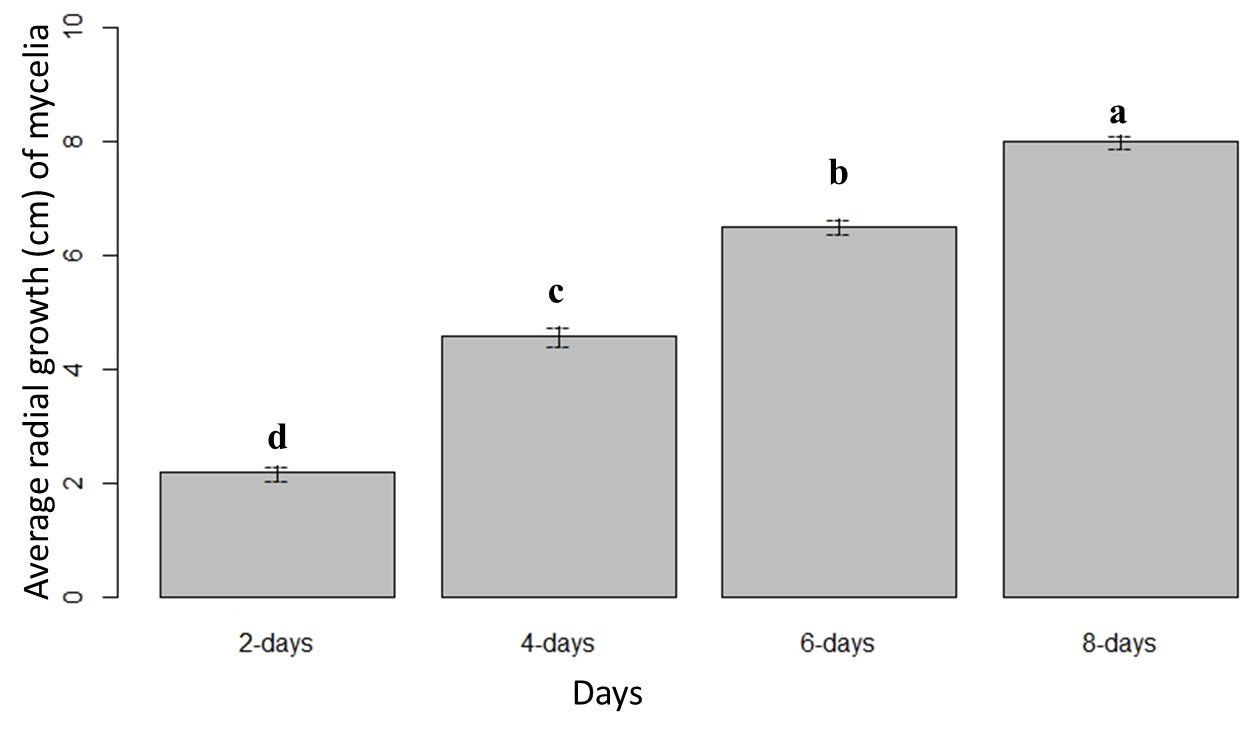
The ANOVA Table 1 showed significant variation between the culture media at α = 0.05, with the least significant difference of 0.006 at 95% confidence level. The highest and lowest mean radial growth were observed in CV8 and WA media, respectively (Fig. 1). The mean radial growth of mycelia was significantly varied over the four different time points. Mycelial growth was fast between the 2 to 4-day period in all media, and later advanced more slowly (Fig. 2). The combined effect (culture media × time points) showed a significant variation on the mycelial growth at 0.001 significance level.
|
|
|
|
Figure 1: Mean radial growth of mycelia on six culture media. Means followed by the same letters are not significantly different at α = 0.05 according to LSD: 0.06, and Mean Square Error = 0.006, at 95% Confidence level. |
Figure 2: Mean radial growth of mycelia over four different time points. Means followed by the same letters are not significantly different at α = 0.05 according to LSD: 0.04, and Mean Square Error = 0.005, at 95% Confidence level. |
Effect of different culture media, and days on the number of sclerotia:
Six different culture media (see Materials and Methods) were used to identify their respective effects on the number of sclerotia. The ANOVA Table 2 showed significant variation between the culture media at α = 0.05, with the least significant difference 3.50 at 95% confidence level.
|
Source of variation |
Df |
Sum Sq |
Mean Sq |
F value |
Pr(>F) |
|
Block |
3 |
176 |
58.8 |
2.722 |
0.08126 |
|
Culture Media |
5 |
88960 |
17791.9 |
823.409 |
< 2e-16 *** |
|
Error a |
15 |
324 |
21.6 |
|
|
|
Time points/Days |
3 |
63375 |
21125.1 |
677.736 |
< 2e-16 *** |
|
Culture Media × Days |
15 |
54064 |
3604.2 |
115.631 |
< 2e-16 *** |
|
Error b |
54 |
1683 |
31.2 |
|
|
|
Significance. Codes: ‘ ’Non-significant, ‘***’ 0.001, ‘**’ 0.01, ‘*’ 0.05. |
|||||
|
cv(a) = 11.9 %, cv(b) = 14.2 %, Mean = 39.21875 |
|||||
Table 2: Analysis of variance for the effect of six culture media on the number of sclerotia (R. solani)
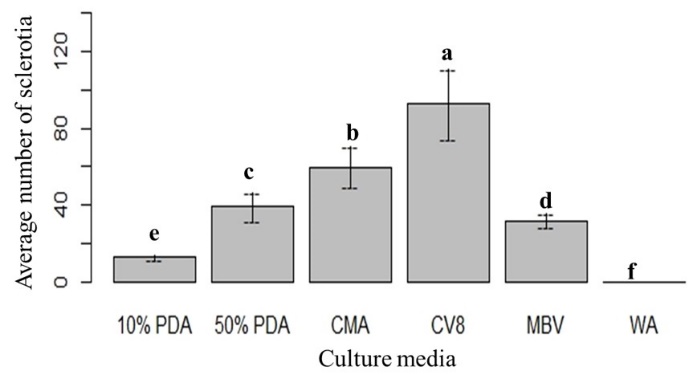
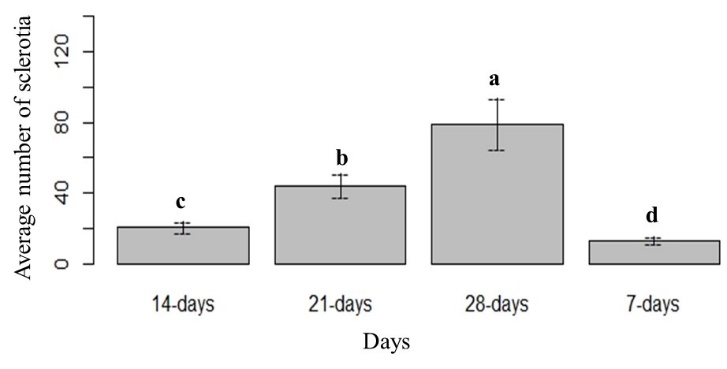
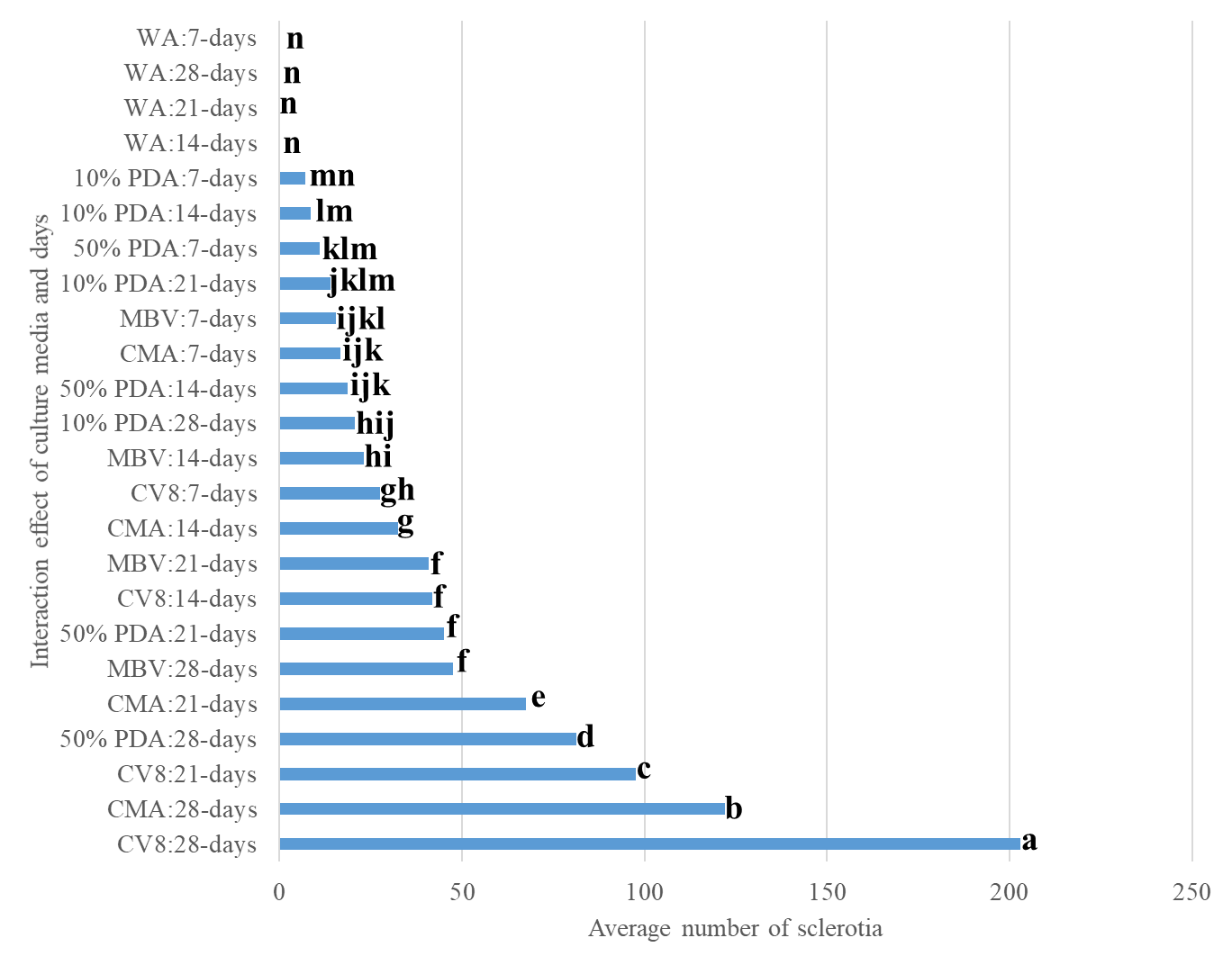
The highest mean number of sclerotia was observed in CV8, followed by CMA. No sclerotia were observed in WA (Fig. 3). The lowest mean number of sclerotia was observed in 10% PDA. The mean number of sclerotia varied significantly over the four different time points. The lowest and maximum number of sclerotia were recorded at 7 days and 28 days, respectively (Fig. 4 and Fig. 5). The interaction effect (culture media × time points/days) showed a significant variation on the mean number of sclerotia, at α = 0.05 according to LSD 7.91 at 95% confidence level (Fig. 6).
|
|
|
|
Figure 3: The mean number of sclerotia on six culture media. Means followed by the same letters are not significantly different at α = 0.05 according to LSD: 3.50, Mean Square Error = 21.6, at 95% Confidence level. |
Figure 4: The mean number of sclerotia over four different time points. Means followed by the same letters are not significantly different at α = 0.05 according to LSD: 3.23, Mean Square Error = 31.17, at 95% Confidence level.
|

Figure 5: Development of sclerotia on six different media at 28-day; A ‘CV8’, B ‘CMA’, C ‘MBV’, D ‘50% PDA’, E ‘10%PDA’ and F ‘WA’.
|
Figure 6: The mean number of sclerotia, interaction effect culture media × Days. Mean followed by the same letters are not significantly different at α = 0.05 according to LSD: 7.91, Mean Square Error =31.17, at 95% Confidence level. |

Figure 7: Measurement of various size of the sclerotia, varied from 0.5 mm to 9.0 mm

Figure 8: Three different categories of sclerotia based on size; “A” Large (≥4.0 mm), “B” Medium (≥2.0 mm), and “C” small (≥0.5 mm).
|
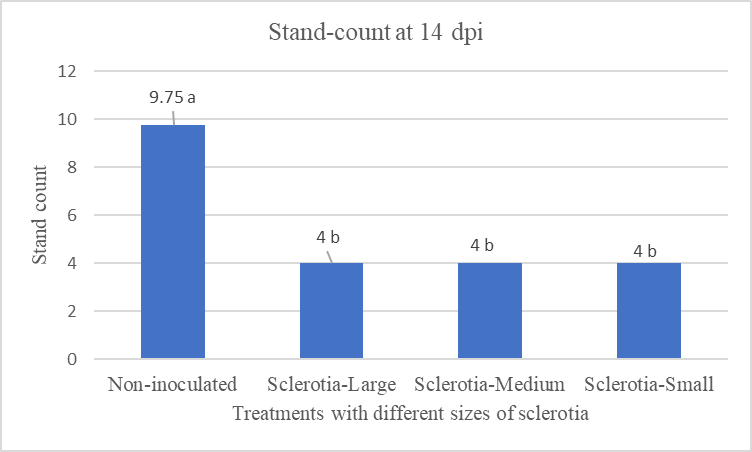
Figure 9: The mean stand count number at 2 weeks of post-inoculation using three different categories of sclerotia. Mean followed by the same letters are not significantly different at α = 0.05 according to LSD: 1.15, Mean Square Error =0.562, at 95% Confidence level. |
Inoculum potentials of different sizes of sclerotia:
To evaluate the size effect of sclerotia on the infection, three different categories of sclerotia were used to inoculate Crystal 101RR (Fig. 7 & 8). The mean stand count number at 14 dpi was statistically similar (Fig. 9). This result suggested that the inoculum potentials (per Garrett, 1956) of different sclerotial sizes used in this investigation did not vary significantly under the conditions of our experiments.
Pre-emergence damping-off studied in vitro using three forms of R. solani inoculum:
In an aseptic technique, three different forms of R. solani inocula demonstrated pre-emergence and post-emergence damping-off at 7dpi, while 100% seedling emergence was observed in the non-inoculated check (Fig. 10). This result demonstrated that three different inoculum types cause similar severity of disease symptoms under in vitro condition.

|
Figure 10: In vitro inoculation on PDA media using three different forms of R. solani inocula; A. plate contained only seed (non-inoculated check), B. plate contained seed and sclerotia, C. plate contained seed and barley inocula, D. plate contained seed and mycelia plug and E. pre-emergence damping-off observed at 7 dpi. |
Assessing Sugar beet Cultivar Sensitivity Under Different Types of Rhizoctonia Inoculum in Greenhouse:
Sugar beet cultivars showed different levels of susceptibility to Rhizoctonia inocula. The mean emergence (or germination) of seven cultivars under three different inoculum types of Rhizoctonia at 14 dpi, was statistically significant (α = 0.05) (ANOVA Table 3).
|
Source of variation |
Df |
Sum Sq |
Mean sq |
F value |
Pr(>F) |
|
Block |
3 |
31 |
10.4 |
0.3175 |
0.81263 |
|
Cultivar |
6 |
3800 |
633.3 |
9.5000 |
8.938e-05 *** |
|
Ea |
18 |
1200 |
66.7 |
2.0323 |
0.02334 * |
|
Rhizoctonia inocula |
3 |
43946 |
14648.5 |
182.5179 |
2.235e-08 *** |
|
Eb |
9 |
722 |
80.3 |
2.4466 |
0.02046 * |
|
Rhizoctonia inocula × Cultivar |
18 |
3086 |
171.4 |
5.2258 |
1.105e-06 *** |
|
Ec |
54 |
1771 |
32.8 |
|
|
|
Significance. Codes: ‘ ’Non-significant, ‘***’ 0.001, ‘**’ 0.01, ‘*’ 0.05. |
|||||
|
cv(a) = 12.7 %, cv(b) = 13.9 %, cv(c) = 8.9 %, Mean = 64.375 |
|||||
Table 3: Analysis of variance for mean emergence of seven cultivars under different forms of Rhizoctonia at 14 dpi
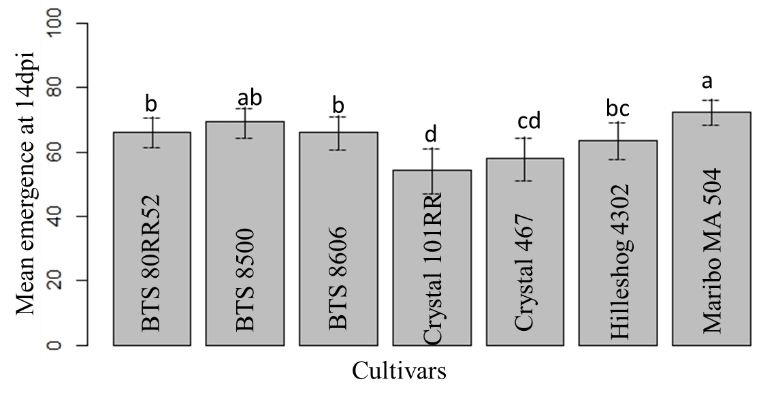
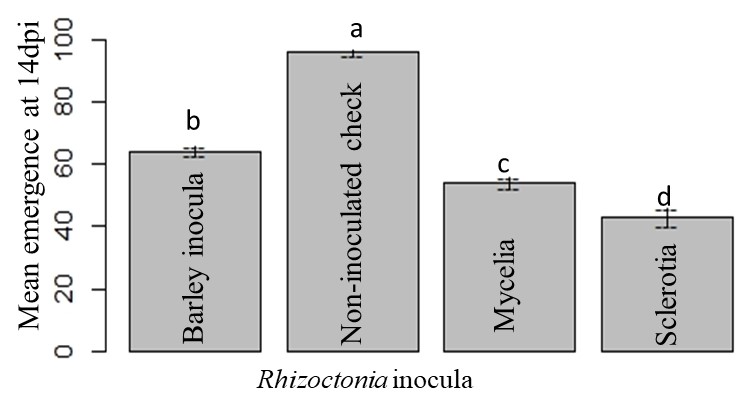
The highest mean emergence was observed in Maribo MA 504, followed by BTS 8500. No significant difference was observed in the mean emergence of BTS 80RR52 and BTS 8600. The lowest mean emergence was found in Crystal 101RR (Fig. 11). Among the three different forms of Rhizoctonia, the lowest mean emergence was observed in sclerotia treatment (Fig. 12). As expected, the non-inoculated check showed the highest mean emergence (Fig. 12).
|
|
|
|
Figure 11: Mean emergence of seven cultivars at 14 dpi. Means followed by the same letters are not significantly different at α = 0.05 according to LSD: 6.06, Mean Square Error = 66.66, at 95% Confidence level. |
Figure 12: Mean emergence under different types of Rhizoctonia inoculation at 14 dpi. Means followed by the same letters are not significantly different at α = 0.05 according to LSD: 5.43, Mean Square Error = 80.25, at 95% Confidence level.
|
|
Source of variation |
Df |
Sum Sq |
Mean sq |
F value |
Pr(>F) |
|
Block |
3 |
71 |
23.8 |
0.7571 |
0.523210 |
|
Cultivar |
6 |
3998 |
666.4 |
10.2864 |
5.330e-05 *** |
|
Ea |
18 |
1166 |
64.8 |
2.0599 |
0.02131 * |
|
Rhizoctonia inocula |
3 |
43729 |
14576.2 |
161.1053 |
3.877e-08 *** |
|
Eb |
9 |
814 |
90.5 |
2.8770 |
0.00754 ** |
|
Rhizoctonia inocula × Cultivar |
18 |
3109 |
172.7 |
5.4921 |
5.345e-07 *** |
|
Ec |
54 |
1698 |
31.4 |
|
|
|
Significance. Codes: ‘ ’Non-significant, ‘***’ 0.001, ‘**’ 0.01, ‘*’ 0.05. |
|||||
|
cv(a) = 22.8 %, cv(b) = 26.9 %, cv(c) = 15.9 %, Mean = 35.35714 |
|||||
Table 4: Analysis of variance for mean pre-emergence damping-off of seven cultivars under a different form of Rhizoctonia inoculation
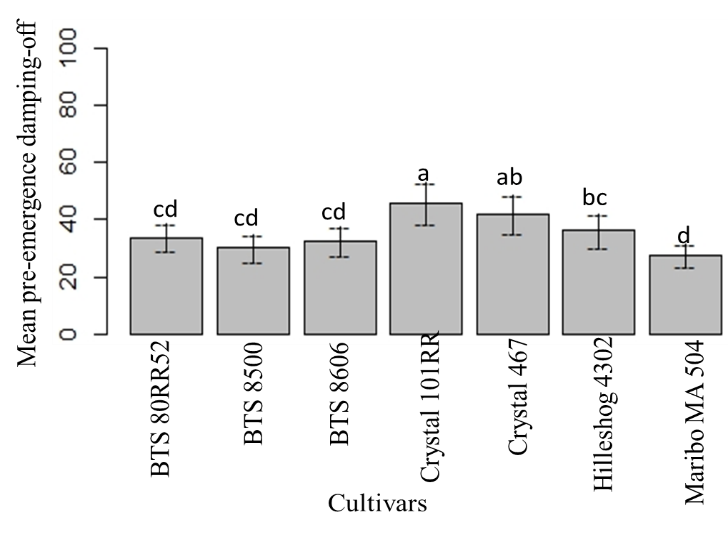
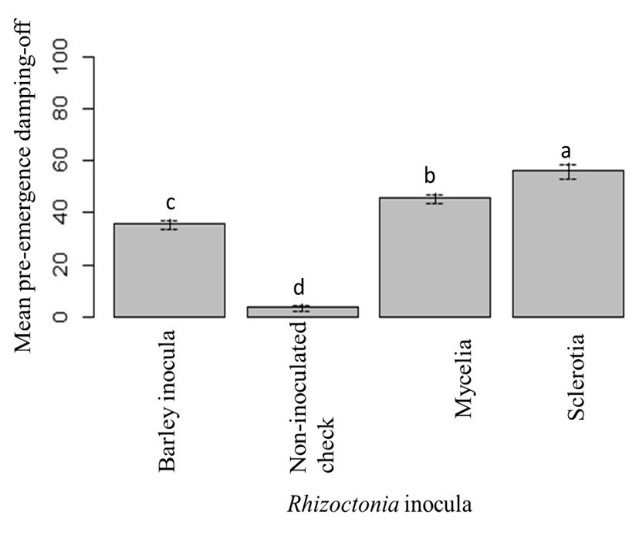
Pre-emergence and post-emergence damping-off was recorded at 2-weeks and 6-weeks post-inoculation, respectively (Yassin 2013). ANOVA generated for pre-emergence damping-off showed statistically significant differences between the cultivars, treatments, and interactions (Table 4). The highest mean pre-emergence damping-off was found in Crystal 101RR, followed by Crystal 467. Mean pre-emergence damping-off of BTS 80RR52, BTS 8500, and BTS 8606 were statistically similar. The lowest mean pre-emergence damping-off was observed in Maribo MA 504 (Fig. 13). Among the three different forms of inocula, the highest pre-emergence damping-off was observed in sclerotial treatment, followed by mycelial treatment (Fig. 14).
|
|
|
|
Figure 13: Mean pre-emergence damping-off of seven cultivars at 14 dpi. Means followed by the same letters are not significantly different at α = 0.05 according to LSD: 7.607, Mean Square Error = 90.47, at 95% Confidence level. |
Figure 14: Mean pre-emergence damping-off under different types of Rhizoctonia inoculation at 14 di. Means followed by the same letters are not significantly different at α = 0.05 according to LSD: 5.75, Mean Square Error = 90.47, at 95% Confidence level. |
The mean post-emergence damping-off was statistically different between the cultivars, treatments, and interactions (ANOVA Table 5).
|
Source of variation |
Df |
Sum Sq |
Mean sq |
F value |
Pr(>F) |
|
Block |
3 |
160.7 |
53.57 |
1.5713 |
0.2069 |
|
Cultivar |
6 |
3483.9 |
580.65 |
11.5901 |
2.401e-05 *** |
|
Ea |
18 |
901.8 |
50.10 |
1.4694 |
0.1386 |
|
Rhizoctonia inocula |
3 |
8610.7 |
2870.24 |
65.1622 |
1.999e-06 *** |
|
Eb |
9 |
396.4 |
44.05 |
1.2919 |
0.2628 |
|
Rhizoctonia inocula × Cultivar |
18 |
4001.8 |
222.32 |
6.5209 |
3.754e-08 *** |
|
Ec |
54 |
1481.1 |
34.09 |
|
|
|
Significance. Codes: ‘ ’Non-significant, ‘***’ 0.001, ‘**’ 0.01, ‘*’ 0.05. |
|||||
|
cv(a) = 47.8 %, cv(b) = 44.8 %, cv(c) = 39.4 %, Mean = 14.82143 |
|||||
Table 5: Analysis of variance for mean post-emergence damping-off of seven cultivars under a different form of Rhizoctonia inoculation
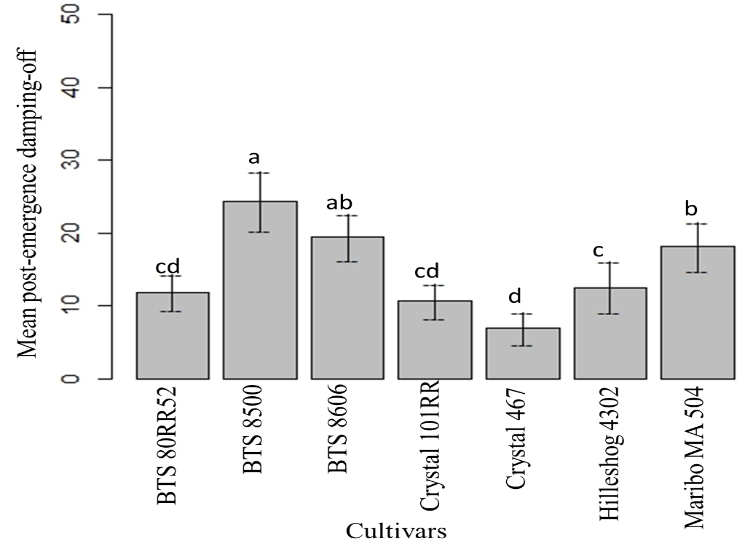
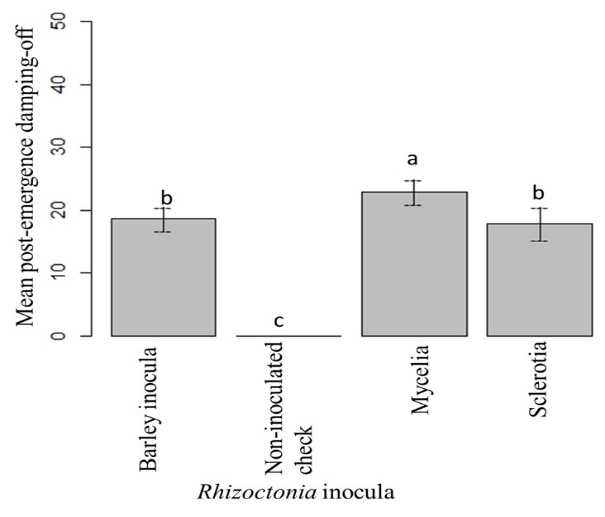
The highest mean post-emergence damping-off was found in BTS 8500, followed by BTS 8606. Mean post-emergence damping-off of BTS 80RR52 and Crystal 101RR were statistically similar. The lowest mean post-emergence damping-off was observed in Crystal 467 (Fig. 15). The highest post-emergence damping-off was observed with mycelial inoculation, followed by sclerotial inoculation. Colonized barley and sclerotia were observed to be statistically similar. There was no post-emergence damping-off in non-inoculated checks of the cultivars (Fig. 16).
|
|
|
|
Figure 15: Mean post-emergence damping-off of seven cultivars up to 42 dpi. Means followed by the same letters are not significantly different at α = 0.05 according to LSD: 5.257, Mean Square Error = 50.09, at 95% Confidence level. |
Figure 16: Mean post-emergence damping-off under different types of Rhizoctonia inoculation up to 42 dpi. Means followed by the same letters are not significantly different at α = 0.05 according to LSD: 4.01, Mean Square Error = 44.04, at 95% Confidence level. |
The mean stand count of the cultivar under three different forms of Rhizoctonia inoculation at 42 dpi was statistically significant. The interactions were found to be significant at α = 0.05. (ANOVA Table 6).
|
Source of variation |
Df |
Sum Sq |
Mean sq |
F value |
Pr(>F) |
|
Block |
3 |
379 |
126.2 |
3.0047 |
0.03822 * |
|
Cultivar |
6 |
1861 |
310.1 |
11.2446 |
2.946e-05 *** |
|
Ea |
18 |
496 |
27.6 |
0.6567 |
0.83654 |
|
Rhizoctonia inocula |
3 |
86193 |
28731.0 |
341.5189 |
1.383e-09 *** |
|
Eb |
9 |
757 |
84.1 |
2.0031 |
0.05671 |
|
Rhizoctonia inocula × Cultivar |
18 |
3032 |
168.5 |
4.0110 |
3.781e-05 *** |
|
Ec |
54 |
2268 |
42. |
|
|
|
Significance. Codes: ‘ ’Non-significant, ‘***’ 0.001, ‘**’ 0.01, ‘*’ 0.05. |
|||||
|
cv(a) = 10.6 %, cv(b) = 18.5 %, cv(c) = 13.1 %, Mean = 49.64286 |
|||||
Table 6: Analysis of variance for the stand count of seven cultivars at 42 dpi
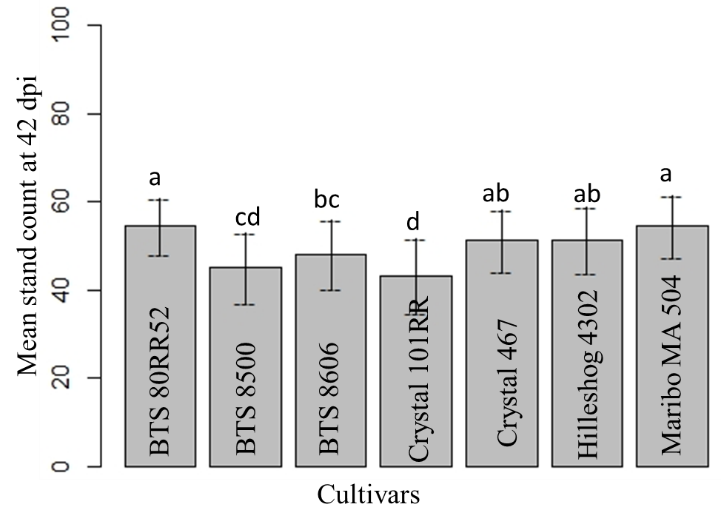
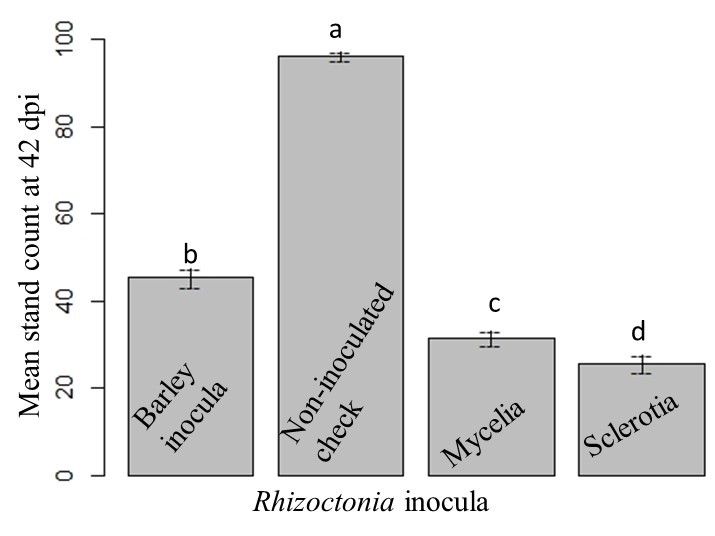
The maximum mean stand count was found in BTS 80RR52 and Maribo MA 504. Mean stand count of Crystal 467 and Hileshog 4302 were statistically similar, followed by BTS 8606. The lowest mean stand count was observed in Crystal 101RR and BTS8500 (Fig. 17). The highest and lowest mean stand counts were observed in the non-inoculated check and sclerotial treatment at 42 dpi, respectively (Fig. 18).
|
|
|
|
Figure 17: Mean stand count at 42 dpi. Means followed by the same letters are not significantly different at α = 0.05 according to LSD: 3.90, Mean Square Error = 27.57, at 95% Confidence level. |
Figure 18: Mean stand count under different types of Rhizoctonia inoculation at 42 dpi. Means followed by the same letters are not significantly different at α = 0.05 according to LSD: 5.54, Mean Square Error = 84.12, at 95% Confidence level. |
The mean root rot ratings of the cultivar under three different forms of Rhizoctonia at 56 dpi were statistically different. The interaction was significant at α = 0.05. (ANOVA Table 7; Fig. 19 and 20).
|
Source of variation |
Df |
Sum Sq |
Mean sq |
F value |
Pr(>F) |
|
Block |
3 |
3.098 |
1.033 |
2.9546 |
0.04054 * |
|
Cultivar |
6 |
26.875 |
4.479 |
9.6681 |
7.983e-05 *** |
|
Ea |
18 |
8.339 |
0.463 |
1.3254 |
0.20980 |
|
Rhizoctonia inocula |
3 |
95.527 |
31.842 |
58.0416 |
3.278e-06 *** |
|
Eb |
9 |
4.938 |
0.549 |
1.5695 |
0.14812 |
|
Rhizoctonia inocula × Cultivar |
18 |
49.911 |
2.773 |
7.9328 |
1.388e-09 *** |
|
Ec |
54 |
18.875 |
0.350 |
|
|
|
Significance. Codes: ‘ ’Non-significant, ‘***’ 0.001, ‘**’ 0.01, ‘*’ 0.05. |
|||||
|
cv(a) = 27.9 %, cv(b) = 30.4 %, cv(c) = 24.3 %, Mean = 2.4375 |
|||||
Table 7: Analysis of variance for disease severity root rot ratings of seven cultivars at 56 dpi
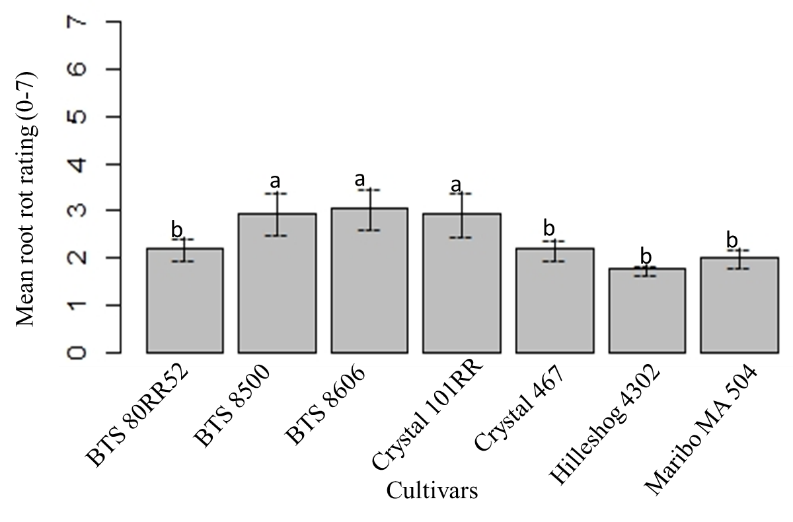
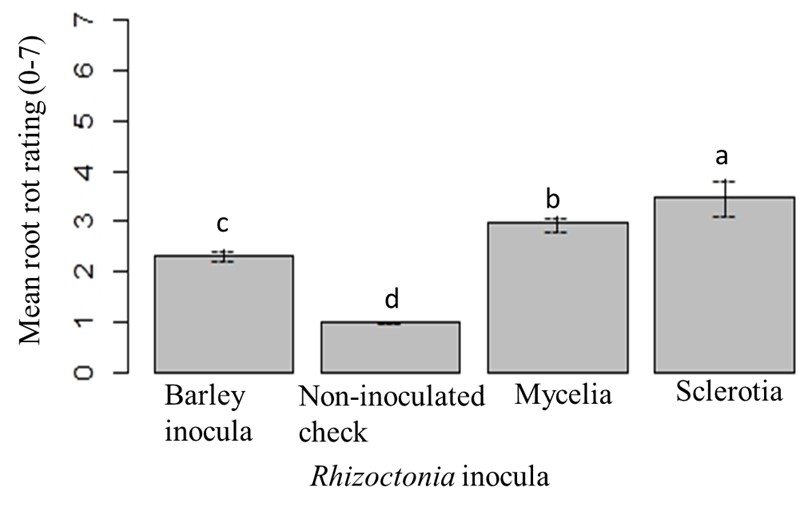
The maximum mean root rot was found in BTS 8500, BTS 8606 and Crystal 101RR. The minimal mean root rot was observed in Hileshog 4302, and statistically similar to BTS 80RR52, Crystal 467 and Maribo MA 504 (Fig. 19). The highest mean root rot was observed with sclerotial inoculum, followed by mycelial inoculum at 56 dpi. No root rot was observed in non-inoculated check (Fig. 20).
|
|
|
|
Figure 19: Mean root rot ratings of seven cultivars at 56 dpi. Mean followed by the same letters are not significantly different at α = 0.05 according to LSD: 0.505, Mean Square Error = 0.463, at 95% Confidence level. |
Figure 20: Mean root rot ratings under different types of Rhizoctonia inoculation at 56 dpi. Mean followed by the same letters are not significantly different at α = 0.05 according to LSD: 0.447, Mean Square Error = 0.548, at 95% Confidence level. |
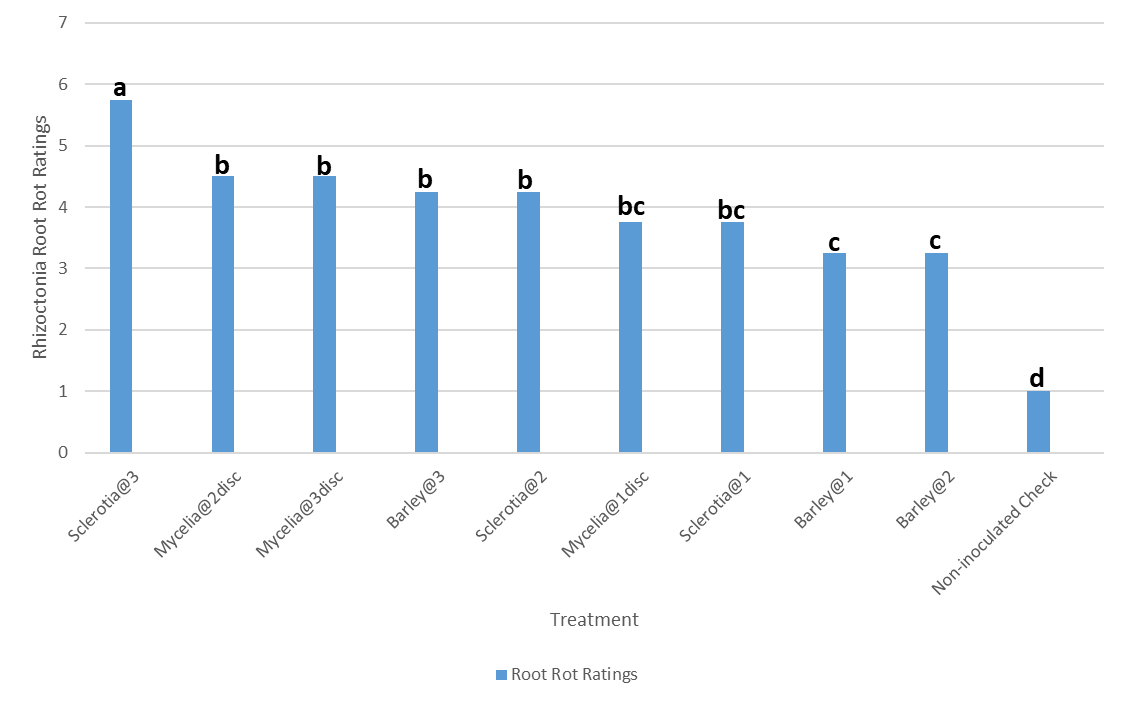
Figure 21: Comparison of Rhizoctonia Root Rot ratings using three different forms and rates of Rhizoctonia inocula in Crystal 101RR in 2019 at Hickson, ND. Means followed by the same letters are not significantly different at α = 0.05 according to LSD: 0.8843, Mean Square Error = 0.375, at 95% Confidence level.
Ten different treatments including the non-inoculated check were used to evaluate the root rot ratings by the three forms of Rhizoctonia inocula in Crystal 101RR. Among the ten treatments, the highest beet root rot was observed with three sclerotia per inoculation. As expected, the non-inoculated check showed healthy seedlings. Two and three mycelial discs/inoculation, as well as two and three barley inocula/inoculation showed moderate effects on root rot. Likewise, one mycelial disc/inoculation and one sclerotium/inoculation showed minimum effects on root rot. One and two colonized barley grains/inoculation showed the lowest root rot ratings (Fig. 21).

Figure 22: Comparison of Rhizoctonia root rot ratings reported from the American Crystal Sugar Company (ACSC) studies with sclerotia-inoculation mediated root rot (1-7 scale) of seven cultivars in greenhouse. Mean damping-off (pre- and post-emergence) of seven cultivars in the bar graph followed by the same letters are not significantly different at α = 0.05 according to LSD: 13.98, Mean Square Error = 90.47, at 95% Confidence level.
Comparison of sclerotia based damping-off, root rot and ACSC commercial ratings:
Sclerotia mediated combined pre- and post-emergence damping-off of seven commercial cultivars significantly varied in the greenhouse study (Fig. 22). The highest mean damping-off was observed in Crystal 101RR, followed by BTS 8606, BTS 8500, Crystal 467RR and Maribo MA 504. BTS 80RR52 and Hilleshog 4302 showed statistically similar mean damping-off. Furthermore, sclerotia mediated root rot in the greenhouse was compared with the commercial (ACSC) root rot ratings of seven cultivars (Source: https://www.sbreb.org/wp-content/uploads/2018/08/NDSU17BW.pdf). There was significant variation between the two types of root rot ratings. In the greenhouse study, the maximum sclerotia mediated root rot was found in Crystal 101RR, which was higher than the commercial root rot. BTS 80RR52 showed similar root rot ratings for both the ACSC reports and the present sclerotial inoculations. In line with the ACSC ratings, the lowest and highest mean root rots with sclerotial inoculation were observed in Hileshog 4302 and Crystal 101RR, respectively (Fig. 22).
Discussion:
Six different culture media were evaluated to determine the effective in vitro methods of large-scale induction of sclerotia in R. solani. The previous study by Hardar et al. (1981) had demonstrated that sclerotia formation of Sclerotium rolfsii is induced in agar media within 72 h following scrapping of aerial mycelia with a scalpel. We, for the first time, demonstrated that amended CV8 is a promising medium to produce Rhizoctonia sclerotia in large scale, without using any abiotic stress. The size of the sclerotia varied significantly among the different culture media but the inoculation potentials of different size sclerotia remained uniformly similar. This experiment showed an easier and more cost-effective way to generate sclerotial propagules as natural inoculum in the laboratory for pathogenic investigations.
R. solani overwinters in soil and crop residues in sugar beet fields, and sclerotia, as well as melanized and moniloid mycelia are the primary source of infection during the seed germination stage (Boland et al. 2004; Lee and Rush 1983). However, earlier studies testing susceptibility to Rhizoctonia were mostly confined to inoculation and in-field screening on adult sugar beets. Those studies used unnatural inocula for pathogenicity trials, mostly consisting of blended Rhizoctonia mycelium or Rhizoctonia-colonized grains. Thus, those screening methods avoided the seed germination stage, which is the most vulnerable to pre- and post-emergence damping off by R. solani. Moreover, we observed an inconsistency in resistance ratings between the commercial (in field) and greenhouse trials of seven cultivars with the three types of inocula. In support of this conclusion, Liu et. al. (2019) demonstrated that most commercially claimed resistant sugar beet cultivars are highly susceptible to Rhizoctonia damping off at the seed germination stage. We, therefore, felt the need to compare and screen the commercial cultivars of known adult plant resistance at the seed germination phase via natural inoculation to figure out reliable resistant ratings of commercial cultivars. This would help identify the best commercial cultivars resistant to R. solani, and further assist sugar beet growers in achieving more economic return.
In vitro inoculation study with three different types of R. solani inocula showed that the pathogenesis of sclerotia and the mycelial plug was better than colonized barley inoculum, causing pre-emergence and post-emergence damping-off. This result demonstrated a novel approach of in vitro inoculation with three different forms of R. solani inocula in PDA media for studying host-pathogen interaction. This study further demonstrated the use of sclerotia or mycelial plugs as a substitute for colonized barley/wheat/millet inoculum for reevaluating the disease ratings of the commercial cultivars, as well as to simulate natural infection in the greenhouse or in-field.
In vivo inoculation studies (greenhouse) showed that all three forms of Rhizoctonia inocula were virulent and capable of damaging sugar beet cultivars. Inoculation with sclerotia showed higher pre-emergence damping-off and root rot in the cultivars compared with the colonized barley. This finding is in line with observations that sclerotia cause seed rot or pre-emergence damping-off as described by other research groups (Gaskill 1968; Naito and Makino 1995). This is the first report evaluating varietal resistance against Rhizoctonia damping-off of commercial cultivars using sclerotia and mycelial plugs. Inoculation with mycelial-plug showed the maximum mean post-emergence damping-off at 42 dpi, followed by sclerotia. The pathogenicity of sclerotia was similar to colonized barley inoculum. These findings suggested that sclerotia and mycelial plugs can be used as a substitute for colonized barley inoculum to evaluate varietal resistance prior to release as a commercial cultivar. Recently, (Liu) et al. (2019) observed that sugar beet cultivars were highly susceptible to R. solani prior to attaining the six- to eight-leaf stage (4-5 weeks) after planting, regardless of the assigned level of resistance. The response of sugar beet cultivars was a major factor in the event of pre-emergence and post-emergence damping-off, root rot disease severity index, and stand count. Maribo MA 504 and BTS 80RR52 showed significantly lower pre-emergence and post-emergence damping-off compared to all other cultivars. Both cultivars showed the highest stand count and lowest root rot. This finding suggested that both Maribo MA504 and BTS 80RR52 can be used as a moderately resistant cultivar group, where there is a minimal incidence of Rhizoctonia root rot. Likewise, Crystal 101RR and Crystal 467 were more susceptible to pre-emergence damping-off but less susceptible to post-emergence damping-off. However, the mean root rot ratings of Crystal 101RR was significantly higher than Crystal 467. This study indicated, Crystal 101RR to be a susceptible cultivar at the seedling phase, therefore, it should not be recommended for cultivation if there is a previous record of Rhizoctonia infestation in the field. BTS 8500, and BTS 8606 were less susceptible to pre-emergence damping-off but they were more susceptible to post-emergence damping-off. Both of them were found to be more susceptible to root rot disease at 56 dpi. This result demonstrated that BTS 8500 and BTS 8606 were susceptible to three different types of Rhizoctonia inocula. However, the growth phase of Crystal 101RR, BTS 8500 and BTS 8606 in areas with any prior history of R. solani can be protected using fungicides, for example, recommended dose of azoxystrobin in-furrow or labeled fungicidal treatment during the early stage, regardless of varietal resistance (Khan 2018).
A field pathogenicity study of three different types of Rhizoctonia inoculum, and its different rates were evaluated at Hickson, ND in 2019. The highest beet root rot was demonstrated with three sclerotial inoculations per plant in Crystal101RR. Two, and three mycelial discs per plant showed moderate root rot, which was similar to two, and three barley inocula. Crystal 101RR was found to be highly susceptible to Rhizoctonia both at the seedling stage with sclerotia inocula (in this investigation), and at the adult phase as reported by the ACSC (Fig. 22). This finding further suggested that sclerotia can be used as natural inoculum in the screening of cultivars for resistance to Rhizoctonia root rot.
In the greenhouse study, sclerotia mediated damping-off (pre- and post-emergence) showed significant variation among the cultivars. This line of evidence further emphasized the need to screen commercially resistant cultivars in the early stage of growth through sclerotia/mycelial plug inoculation to get a more reliable resistance rating. Although growers in North America and Europe commonly use effective fungicides such as sedaxane or seeds coated with fungicides to control pre-emergence damping-off (Liu and Khan 2016), but yet 5-10% seedlings in an acreage fall due to pre- and post-emergence damping-off. Moreover, the seed companies need to consider age-dependent inoculation of plants for better evaluation of cultivars against susceptibility to R. solani. Furthermore, sclerotia mediated root rot ratings in the greenhouse largely differed from the commercial (ACSC) root rot ratings of the seven cultivars. This finding suggested a need to reevaluate the resistance of the commercial cultivars based on natural inoculum in the greenhouse.
In conclusion, R. solani is a devastating pathogen causing economic losses in sugar beet production. The disease pressure is escalating in Europe and the USA. An understanding of Rhizoctonia virulence with natural inocula (sclerotia or mycelia) and screening of cultivars at the seed germination stage are imperative for successful disease management. Future studies should look into interactions of cultivars at the seedling stage with other anastomosis groups of R. solani in order to minimize stand and yield loss in sugar beets.
Acknowledgments:
Authors are very much thankful to D. Lakshman (USDA, ARS, Baltimore, Beltsville, MD 207052-350), Aiming Qi (School of Life and Medical Sciences, University of Hertfordshire, Hatfield, AL10 9AB, UK) and M. F. R. Khan (Department of Plant Pathology, North Dakota State University, Fargo, ND 58102, University of Minnesota, St. Paul, MN USA) for their suggestions and technical support.
Conflict of Interest Statement: Authors declared no conflict of interest

Open Access By Aditum Open Access Journals id licensed under Creative Commons Attribution 4.0 International License. Based On a Work at aditum.org