
Samim Ali1, Satyawart Sangwan2, Priyam Srivastava3, Shahzeb Khan4, Vikas Raghuvanshi5, Amarjeet Kumar5, Ankur Nawani6, Vishal Chandra3 and Pramod Yadav3,6*
1ICMR- National Institute of Cancer Prevention and Research, Noida, Uttar Pradesh, India.
2Kalpana Chawla Government Medical College, Karnal, Haryana, India.
3School of Life Sciences and Biotechnology, Chhatrapati Shahu Ji Maharaj University Kanpur, Uttar Pradesh, India.
4Department of Biotechnology, Himachal Pradesh University, Shimla, India.
5School of Biotechnology, Madurai Kamaraj University, Tamil Nadu, India.
6Amity Institute of Neuropsychology and Neurosciences, Amity University Uttar Pradesh, India.
*Corresponding Author: Pramod Yadav, Amity Institute of Neuropsychology and Neurosciences, Amity University Uttar Pradesh, India.
Received Date: April 29, 2024
Accepted Date: May 06, 2024
Published Date: May 10, 2024
Citation: Samim Ali, Satyawart Sangwan, Priyam Srivastava, Shahzeb Khan and Pramod Yadav.et.al. (2024) “From Bronchial Asthma to Tuberculosis: Unravelling the Cytokine-Macrophage Axis in Diverse Diseases.”, International Journal of Medical Case Reports and Medical Research, 2(5); DOI: 10.61148/2994-6905/IJMCRMR/041.
Copyright: © 2024. Pramod Yadav. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Cytokines, tiny proteins secreted by cells, play a critical role in mediating communication and interactions between cells. They also function as immunomodulating agents, adjusting immune responses. When released into the bloodstream or tissues, cytokines bind to specific receptors on target immune cells, triggering cellular responses. Cytokines are implicated in various diseases, including asthma, COPD, HIV infection, and multiple sclerosis. This review delves into the intricate interplay between cytokines and macrophages, focusing on their roles in inflammation and the immune response. Macrophages, which are scavenger cells of the immune system, exhibit remarkable heterogeneity, reside in diverse tissues, and play crucial roles in both innate and acquired immunity. They can be activated for proinflammatory or anti-inflammatory functions, contributing to tissue destruction or regeneration. Cytokines influence macrophage activation and polarization, impacting the inflammatory process. Dysregulated cytokine production and macrophage activation are observed in various diseases. For example, in asthma, an imbalance in macrophage phenotypes contributes to airway hyperresponsiveness. Understanding the complex interplay between cytokines and macrophages is crucial for developing novel therapeutic strategies for inflammatory diseases. Future research directions include utilizing humanized animal models, single-cell sequencing, and immunomodulatory therapies to further decipher the intricate roles of cytokines and macrophages in health and disease. Ultimately, elucidating these interactions holds immense potential for improving human health outcomes.
1. Introduction:
Cytokines, proteins secreted by cells, play pivotal roles in mediating intercellular communication and interactions. These proteins are also referred to as “lymphokines,” “monokines,” “chemokines,” and “interleukins” (when produced by one leukocyte and acting on other leukocytes). The term ‘cytokine’ originates from the Greek words ‘cyto’ (cell) and ‘kinos’ (movement) (Kany et al., 2019a). Recently, cytokines have been referred to as ‘immunomodulatory agents’ due to their ability to adjust immune responses. Cytokines are released into the bloodstream or directly into tissues, where they bind to their target immune cell receptors, triggering specific cellular responses. Inflammatory pathogen-associated molecular patterns (PAMPs), such as heat shock proteins (HSPs), peptidoglycans (PGNs), and lipopolysaccharides (LPS), as well as DAMPs, such as HMGB1 and adenosine triphosphate, are key players in this process (Kany et al., 2019a). These molecules, originally intracellular proteins or nucleic acids are not typically found outside the cell. Pattern recognition receptors (PRRs) recognize PAMPs and DAMPs. The primary PRRs involved in inflammatory pathways are TLR, NLR, and MBL (Kany et al., 2019a). Upon engagement, PRRs transmit signals within the cell, for instance, through MAP kinase signaling pathways to the nucleus, where various transcription factors are activated. Additionally, two polypeptide chains are present: a cytokine-specific α subunit and a signal-transducing β subunit. Upon binding to these receptors, cytokines can induce cellular and humoral immune responses and inflammatory responses, regulate hematopoiesis, control cell proliferation and differentiation, and promote wound healing (Lv et al., 2023).
In the context of bronchial asthma, research has indicated that cytokine production by T cells, rather than the eosinophil concentration or IgE synthesis, is typically associated with altered airway behaviour (Kany et al., 2019a). An increase in the number of CD4+ Th cells of the Th2 subtype has been observed in the airways. In chronic obstructive pulmonary disease (COPD), there is increased expression of IL-4 in bronchoalveolar lavage (BAL) fluid from patients, which is pivotal in the immune response and influences the differentiation of Th0 cells into Th2 cells, which could trigger allergen sensitivity (Kedzierska & Crowe, 2001). IFN-γ, a key cytokine in patient inflammation, facilitates the lung infiltration of Th1 and Tc cells by enhancing the expression of the chemokine receptor CXCR3 on these cells and promoting the secretion of CXCR3-activating chemokines such as CCL9, CCL10, and CCL11 (Gulati et al., n.d.). In the context of HIV infection, cytokines play a vital role in maintaining immune system homeostasis. There was a marked decrease in the release of Th1 cytokines, including IL-2 and IFN-γ, concurrent with an increase in the production of Th2 cytokines (IL-4 and IL-10) and proinflammatory cytokines (IL-1, IL-6, IL-8, and TNF-α) during HIV infection (Gulati et al., n.d.). Moreover, cytokines such as TNF-α, TNF-β, IL-1, and IL-6 have been found to stimulate HIV replication in lymphocytes and monocyte-derived macrophages (MDMs). In the case of multiple sclerosis (MS), neurodegeneration is a direct result of demyelination, leading to the formation of plaques in the white matter, a hallmark pathology of the disease. The cytokines IL-6 and IFN-γ instigate an inflammatory response in the brain’s white matter, contributing to plaque development (Gulati et al., n.d.). Notably, the administration of IFN-γ exacerbates MS symptoms. However, IFN-γ treatment has demonstrated some efficacy in reducing the relapse rate among MS patients.
Considering the pivotal role of cytokines in these diseases, as corroborated by the referenced data and articles, it is essential to further explore the therapeutic potential of cytokines. Consequently, this study aimed to elucidate the role of cytokines and macrophages in several of the most common diseases. A comprehensive literature search was conducted using a variety of keywords, resulting in a plethora of articles. The most pertinent and relevant articles were meticulously selected for this systematic review.
2. Macrophage heterogeneity and functional diversity:
2.1. Macrophages:
Macrophages, which are sentinels of the immune system, exhibit remarkable tissue tropism and strategically position themselves in lymphoid organs, mucosal interfaces, and other pivotal sites. These versatile cells act as resident custodians, wielding their phagocytic ability to maintain tissue homeostasis (Chung, 2005). These cells diligently scavenge apoptotic and necrotic debris while simultaneously standing guard against invading pathogens. In addition to being mere scavengers, macrophages are key players in both innate and adaptive immunity (Chung, 2005). The innate arsenal of these viruses includes immediate neutralization of foreign microorganisms and orchestration of leukocyte recruitment. These proteins meticulously regulate the delicate balance between antigen presentation and clearance through phagocytosis and subsequent degradation. This intricate interplay with T and B lymphocytes, facilitated by a diverse repertoire of cytokines, chemokines, and bioactive molecules, dictates the trajectory of the immune response (Osuji et al., 2018). Macrophage activation, a dynamic process influenced by contextual cues, can skew toward proinflammatory or anti-inflammatory functions. This versatility allows them to orchestrate tissue destruction during infection, followed by a switch toward regenerative and wound healing processes. Ultimately, macrophages act as crucial orchestrators, initiating, instructing, and even terminating the adaptive immune response, ensuring delicately balanced and dynamic defenses against diverse immunological challenges (Catalfamo et al., 2012).
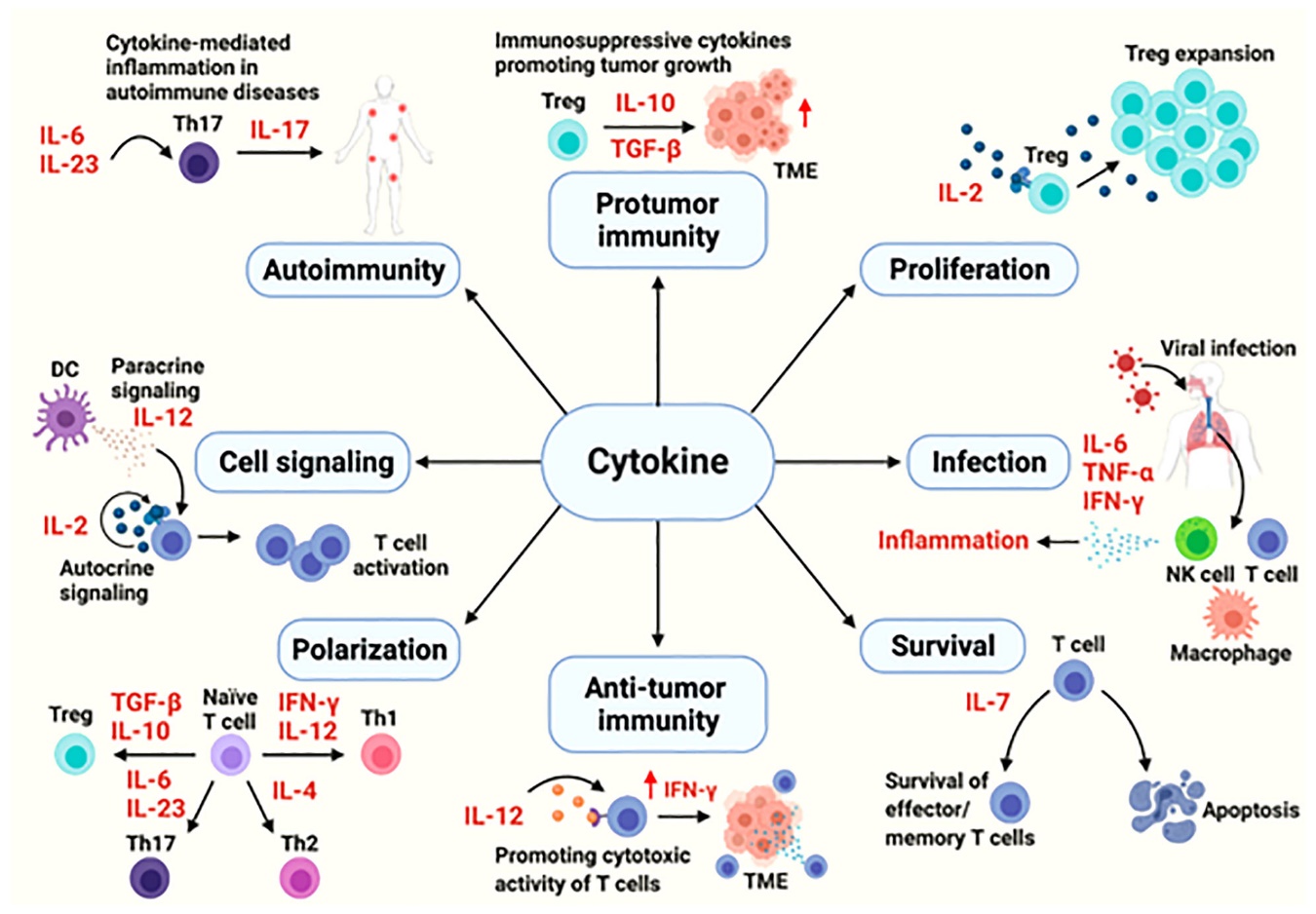
Figure 1: The cytokines in immune processes. [source: (Kang et al., 2021)].
2.2 Classically activated macrophages:
M1 macrophage polarization is orchestrated by the coordinated symphony of IFN-γ and microbial stimuli, particularly LPS. IFN-γ, the lone maestro of type II IFNs, interacts with a dedicated orchestrator conductor, IFNGR, a heterodimer formed by two ligand-capturing IFNGR1 subunits and two signal-transducing IFNGR2 subunits. This exquisite interplay triggers a cascade of molecular events directing macrophage differentiation toward the robust effector repertoire of M1 macrophages (Orecchioni et al., 2019).
2.3. Tumor-associated macrophages:
Environmental cues within the TME orchestrate intricate leukocyte infiltration, proliferation, and polarization, thereby dictating the functional repertoire of recruited macrophages. Among these, TAMs exhibit a protumorigenic phenotype and thus harbor a potent arsenal of proangiogenic and tumor-promoting chemokines, such as CCL2/MCP-1. Hypoxia, a defining hallmark of the TME, further sculpts TAM functionality, igniting a proangiogenic program that amplifies tumor-driven vascularization in an indirect yet potent manner (Xie et al., 2016). However, not all macrophage contributions favor tumor progression. In a murine model, tumor-derived GM-CSF induced macrophage-mediated degradation of the extracellular matrix via upregulation of MMPs, concurrently stimulating angiostatin production, ultimately leading to suppressed metastatic growth (Xia et al., 2023).
2.4. Role of cytokines and macrophages in inflammation:
The intricate nature of inflammation relies on the delicate balance between proinflammatory and resolving signals. Disruption of this equilibrium leads to uncontrolled inflammation, causing cellular and tissue damage (Romero et al., 2007). Macrophages, the stalwart foot soldiers of the mononuclear phagocyte system, orchestrate this intricate dance, playing a pivotal role in initiation, maintenance, and resolution. Elie Metchnikoff, a Nobel laureate, initially christened these phagocytic warriors as "white blood cells" for their frontline defenses against infection. In 1924, Aschoff refined the nomenclature, coining the term "macrophage" for this diverse lineage encompassing monocytes, macrophages, and histiocytes. During inflammation, macrophages carry out three essential steps: antigen presentation, phagocytosis, and immunomodulation through cytokine and growth factor orchestration (Soehnlein & Libby, 2021). Activated by a symphony of signals, including cytokines such as IFN-γ, GM-CSF, and TNF-α; bacterial lipopolysaccharide; and matrix cues, macrophages ignite the inflammatory response. Resolution, the counterpoint to inflammation, necessitates the silencing of these inflammatory conductors and their effector cells. Deactivation of macrophages, achieved through diverse mechanisms, allows tissue repair and restoration (L. Chen et al., 2018). Neutrophilic granulocytes, the rapid responders of the acute inflammatory phase, extravasate through a fleeting set of endothelial adhesion molecules orchestrated by cytokine-induced upregulation. Intriguingly, the duration of inflammation appears to differ between preterm deliveries and term deliveries, with elevated proinflammatory cytokine and proteinase levels preceding cervical dilation. Similarly, chorioamnionitis patients exhibit elevated proinflammatory cytokines across various compartments, potentially contributing to preterm contractions . The role of inflammation in atherosclerosis is a captivating research avenue. Recent studies have investigated (Watanabe et al., 2019)ate interplay between proinflammatory cytokines such as IL-1, IL-18, and OPN and their anti-inflammatory counterparts, including IL-1 receptor antagonists, IL-10, and IL-18-binding proteins (Xia et al., 2023). Additionally, the contribution of chronic infections such as Helicobacter pylori and Chlamydophila pneumoniae to persistent inflammation has garnered significant attention.
2.5. Cytokine therapeutic uses:
IL-1 and IL-2 have demonstrated potential as consistent immunostimulants for combating AIDS. Empirical studies have substantiated the hypothesis that immunostimulatory cytokines can counteract the immunosuppressive characteristics of cancer. The synthesis of IL-10, TGF-β, and PGE2 by ovarian cancer and Cap (3) cells contributes to the comprehensive inhibition of antitumour activities (Soehnlein & Libby, 2021). IL-10 facilitates the differentiation of monocytes into mature macrophages and impedes their differentiation into dendritic cells. The alarming inflammatory response of humans to cytokines such as IL-1, IL-2, IL-3, IL-4, IL-6, IL-12, and TNF-α has resulted in adverse effects. The first cytokine approved for cancer treatment was IL-2, but its proinflammatory effects are poorly tolerated, thereby limiting its efficacy in conditions such as melanoma and renal cell carcinoma (Gulati et al., n.d.). IL-10 has emerged as a promising candidate for various autoimmune diseases because it not only suppresses the production of IFN-γ, IL-1, TNF-α, and IL-6 but also has other anti-inflammatory effects (Gulati et al., n.d.). However, several trials of recombinant human IL-10 have demonstrated limited effectiveness in treating psoriasis, rheumatoid arthritis, and Crohn’s disease, and this cytokine has not been approved for therapeutic use (Dinarello, 2007). The FDA has approved the cytokines IL-2 and IFN-α, with IL-2 being used for the treatment of metastatic melanoma and renal cell carcinoma in high-dose boluses and IFN-α serving as an adjuvant treatment for stage III melanoma, hematologic malignancies, AIDS-related Kaposi’s sarcoma, and in combination with bevacizumab as an antiangiogenic agent for advanced renal cancer (Gulati et al., n.d.). Recently, various cytokines, including IL-7, IL-12, IL-15, IL-18, IL-21, and GM-CSF, have been tested in clinical trials for advanced cancer (Gulati et al., n.d.).
3. Dysregulated Immunity: Cytokines and Macrophage Links
3.1. Macrophages in acute lung disease:
Acute respiratory distress syndrome (ARDS) is a critical condition characterized by the rapid onset of diffuse alveolar damage and impaired gas exchange (Kim & Yang, 2019). Diverse primary and secondary mechanisms can trigger ARDS, including shock, severe sepsis, pulmonary contusion, gastroesophageal reflux, pneumonia, drug toxicity, transfusion, and acute pancreatitis (Arora et al., 2018). Iatrogenic or physical injury from mechanical ventilation can further contribute to ARDS development. The pathogenesis of this disease commences with the rupture of alveolar septa, disruption of the epithelium‒capillary interface, release of protein-rich fluid, secretion of proinflammatory chemokines and cytokines, and infiltration of inflammatory cells such as monocytes and neutrophils. ALI is characterized by an amplified M1 response and impaired M2-mediated repair, while chronic diseases such as fibrosis and cancer are dominated by hyperresponsive, alternatively activated M2 cells (Xie et al., 2016).
3.2 Macrophages in various infections:
During the initial phase of bacterial infection, macrophages exhibit an M1 phenotype. Delayed regulation of macrophage-mediated inflammation results in a cytokine storm, contributing to the pathogenesis of severe sepsis (Xia et al., 2023).During inflammatory responses, macrophages exhibit two key strategies for resolution: programmed cell death (apoptosis) or phenotypic reprogramming toward an M2 state. These mechanisms safeguard the host from excessive tissue damage and initiate the critical transition from inflammation to tissue repair. Apoptosis eliminates potentially harmful proinflammatory macrophages, while the M2 phenotype promotes debris clearance, angiogenesis, and collagen deposition, ultimately facilitating wound healing (Orecchioni et al., 2019).
3.3. Macrophage counts in bronchiectasis:
In bronchiectasis, a chronic pulmonary pathology characterized by persistent inflammation, bacterial colonization, and specific commensal bacteria, such as Stromatosus mucilaginosus, exacerbates disease progression. S. mucilaginosus facilitates the establishment of opportunistic pathogens, notably Pseudomonas aeruginosa, by fostering a conducive microenvironment within compromised airways (Vidaillac & Chotirmall, 2021). This bacterium specifically activates Toll-like receptor 2 (TLR2) on host immune cells, particularly M1 macrophages, triggering a proinflammatory cytokine cascade that perpetuates tissue damage and infection susceptibility (Romero et al., 2007).
3.4. Macrophages in asthma:
Asthmatic inflammation hinges on a precarious balance between proinflammatory and anti-inflammatory pulmonary macrophage functions. While M2 macrophages, champions of tissue repair and lung microenvironment homeostasis, normally serve as allies, their dysregulation unleashes a cascade of pathological events. This dysregulation manifests as airway hyperresponsiveness, fuelled by augmented cell recruitment and mucus hypersecretion, potentially orchestrated by aberrant miRNA expression (Duan et al., 2023).
Recent investigations have shed light on the intriguing nexus between specific miRNAs and alleviating asthmatic symptoms. For instance, miR-145 exhibits therapeutic potential by dampening the expression of proinflammatory cytokines (IL-13 and IL-5), thereby curbing inflammation (Girodet et al., 2016). Similarly, miR-146a orchestrates multiple processes by suppressing an array of proinflammatory cytokines and chemokines, effectively eliminating the inflammatory storm. Furthermore, miR-21 strategically neutralizes IL-21, a pivotal player in Th1 cell polarization, by disrupting a key pathogenic pathway. The intricate link between asthma and miRNAs is further underscored by the observation that let-7 downregulation amplifies the levels of IL-13, a pivotal orchestrator of allergic responses, potentially influencing Th2 skewing (Habib et al., 2022).
3.5. Macrophages in chronic obstructive pulmonary disease (COPD):
Progressive inflammatory and structural derangements orchestrate the pathogenesis of COPD. Chronically inhaled noxious stimuli trigger diverse responses in lung epithelial cells, culminating in a maladaptive cascade (Wang et al., 2018). Epithelial senescence accelerates, the pulmonary capillary vasculature undergoes progressive destruction, and airway remodelling ensues. This architectural disarray manifests as decreased lung compliance, the hallmark functional signature of COPD. The inflammatory cascade in COPD involves a repertoire of key mediators. Cytokines such as TNF-α, IL-1β, and GM-CSF act in both autocrine and paracrine fashions, perpetuating the inflammatory milieu (McCormick et al., 2021). TGFβ orchestrates the nefarious transformation of fibroblasts into myofibroblasts, fueling the fibrotic crescendo of airway remodelling. Recent insights implicate miRNA dysregulation in this macabre dance, with downregulation of miR-152 liberating MMP12, a potent maestro of emphysematous destruction (Birch et al., 2023).
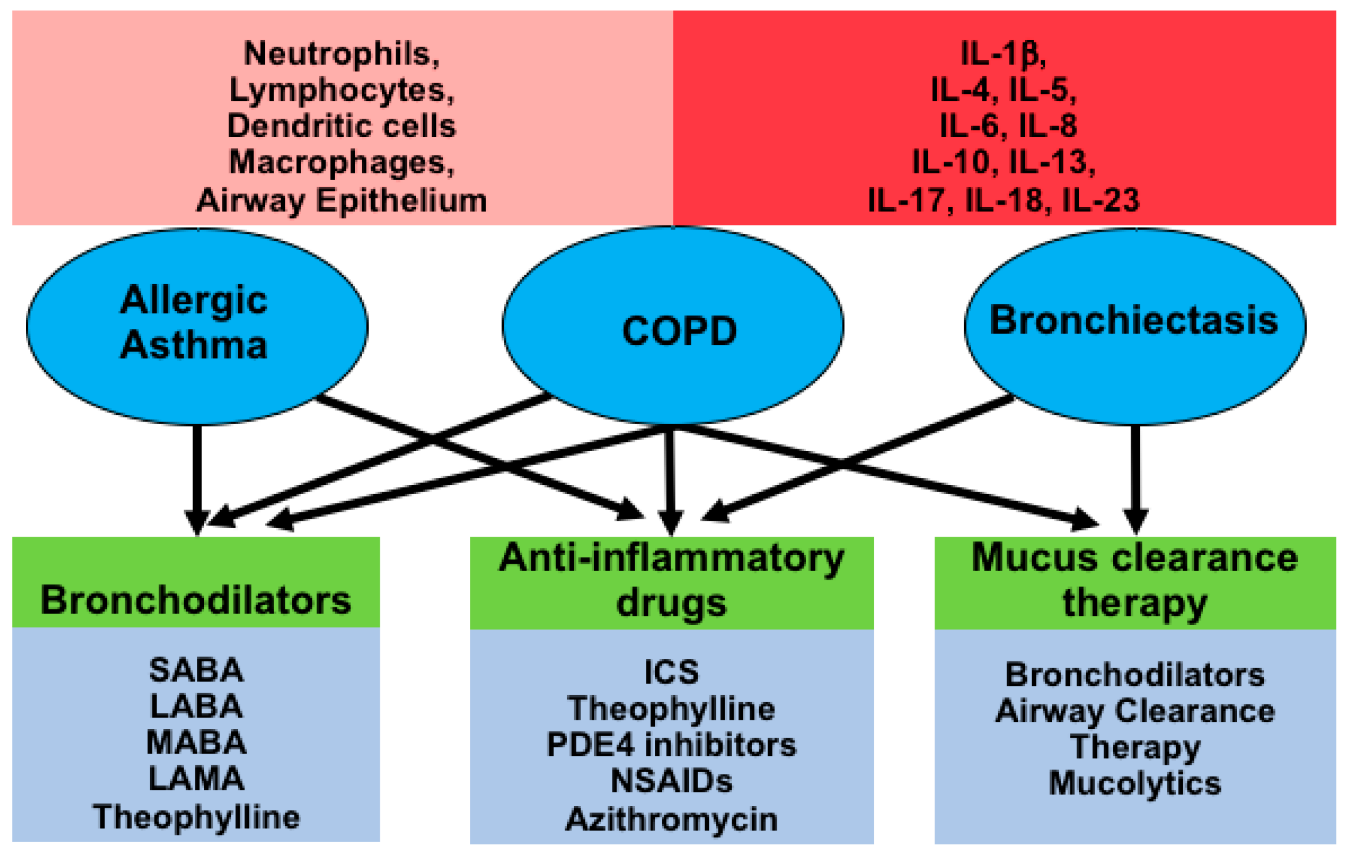
Figure 2: The diagram is summarizing the three chronic inflammatory airway diseases. [source: (Garth et al., 2018)]
3.6. Macrophages in tuberculosis (TB):
Mycobacterium tuberculosis (Mtb), the causative agent of the ubiquitous infectious disease tuberculosis (TB), faces its initial barrier within the alveoli. Alveolar macrophages, the sentinels of pulmonary immunity, engulf Mtb droplets via phagocytosis. Within the phagolysosome, a potent arsenal of reactive oxygen species (ROS) unleashes oxidative mayhem upon invaders (Yadav, 2023). This initial skirmish triggers the recruitment of mononuclear leukocytes, bolstering the immune response. These professional phagocytes constitute the first line of defense, actively engaging and eliminating mycobacterial threats. Among these leukocytes, classically activated macrophages (M1) are central to combating intracellular parasites (Sampath et al., 2018). Their potent microbicidal arsenal, fueled by enhanced antigen presentation and inflammatory cytokine production, spearheads the antimycobacterial response. Alternatively, activated M2 macrophages adopt a different paradigm. These versatile cells prioritize tissue repair, suppress inflammation, and promote wound healing, contributing to a more orchestrated immune response. This dynamic interplay between M1 and M2 macrophages underscores the complex immunological landscape of TB. Understanding these intricate orchestrations paves the way for novel therapeutic strategies aimed at tipping the balance toward pathogen eradication and tissue preservation (Lavalett et al., 2020).
3.7. Kidney diseases:
In chronic kidney disease (CKD), infiltrating immune cells transcend passive bystanders, morphing into detrimental actors within the pathogenic orchestration(Stenvinkel et al., 2021). These inflammatory elements actively propel disease progression, orchestrating a nefarious symphony of nephron attrition and fibrotic encasement. This insidious immune-mediated assault elevates CKD to the ominous ranks of chronic inflammatory diseases, demanding novel therapeutic strategies that not only eliminate inflammation but also disarm the nefarious orchestrators of renal devastation (Fattah et al., 2019). Anti-inflammatory strategies have emerged as common therapeutic targets for renal disorders, given that patients with early-stage CKD exhibit subclinical inflammation and activation of circulating immune cells. Recent investigations have illuminated the versatility and complexity of immune cell roles (Cantero-Navarro et al., 2021). Monocytes/macrophages, a critical type of immune cell, are innate immune system phagocytes found across various organs.
3.8. Autoimmune disease:
Interleukin-6 (IL-6) is a pleiotropic cytokine that critically influences diverse pathological processes, encompassing autoimmune disorders, bacterial infections, and metabolic dysregulations. Composed of four α-helices, this 184-amino acid protein has multiple functions, transcending its initial characterization as a B-cell stimulatory factor (Kishimoto et al., 1986). Notably, IL-6 has potent immunomodulatory effects, promoting CD4+ T-cell expansion via IL-21 induction and guiding CD4+ T-cell differentiation toward the Th2 and Th17 lineages (Pramod et al., 2023; Raghuvanshi et al., 2023; Yadav et al., 2023; Yadav & Yadav, 2023).
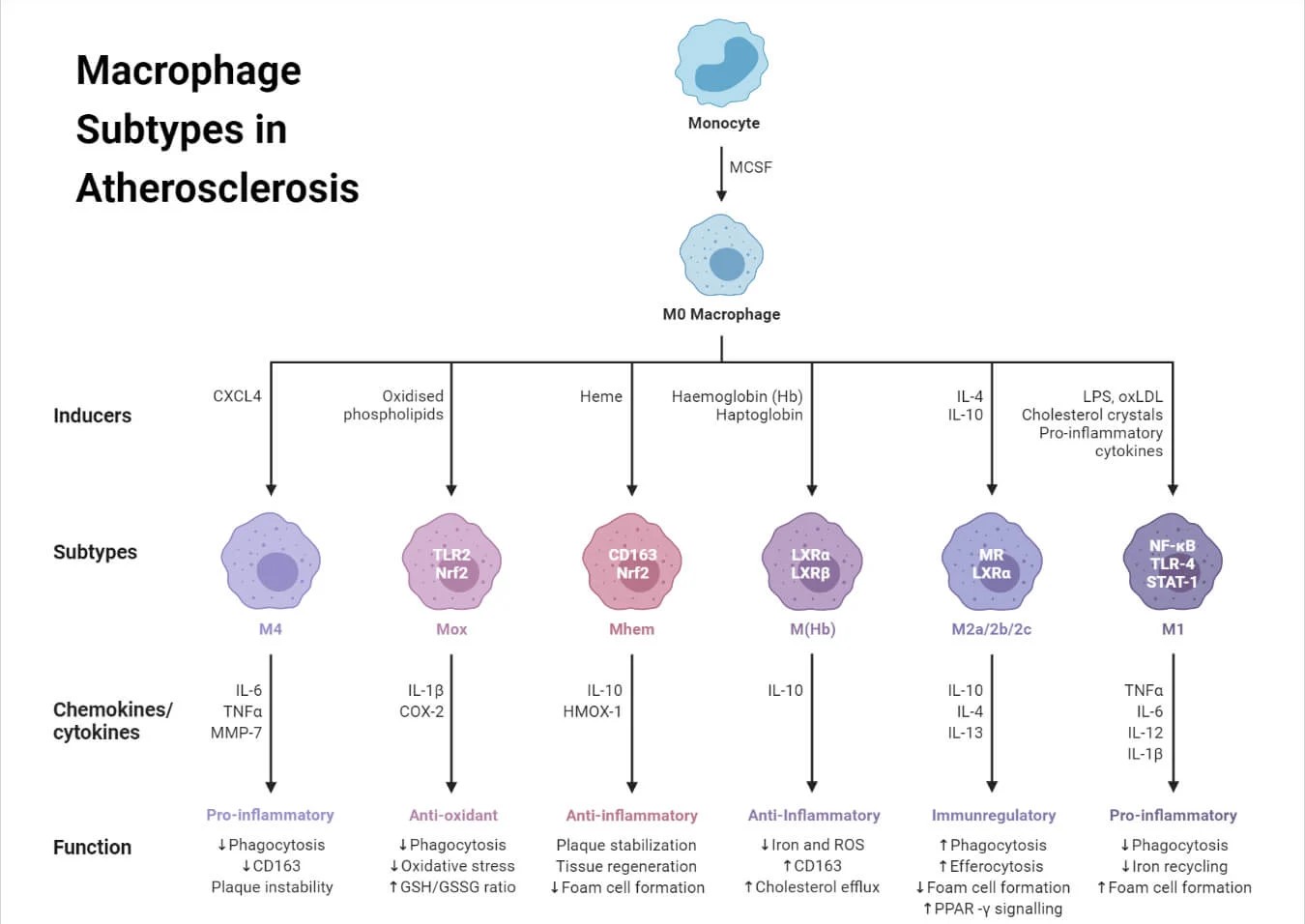
Figure 3: Macrophage Subtypes in Atherosclerosis. [source: (Macrophages: Structure, Immunity, Types, Functions, n.d.)]
3.9. Cardiovascular disease:
Cytokines and macrophages are instrumental in the onset and progression of CVD. Cytokines, signaling molecules produced by various cells, including immune cells such as macrophages, regulate immune responses, inflammation, and cell growth and differentiation (Amin et al., 2020; Haybar et al., 2023). In CVD, cytokines such as TNF-alpha, IL-6, and IL-1beta contribute to the development of atherosclerosis, a primary cause of CVD. Macrophages, a type of immune cell, play a pivotal role in atherosclerosis. In its early stages, macrophages infiltrate the arterial wall and take up modified lipids such as oxidized LDL to form foam cells. These foam cells contribute to the formation of fatty streaks, the earliest visible signs of atherosclerosis (Lv et al., 2023). Macrophages also secrete cytokines and other inflammatory mediators that promote inflammation and atherosclerosis progression. In advanced atherosclerosis, macrophages can form a necrotic core by undergoing apoptosis and releasing their contents, including cholesterol and proinflammatory cytokines. This can lead to plaque instability and rupture, triggering acute cardiovascular events such as myocardial infarction or stroke (Wei et al., 2023).
3.10. Cancer:
In cancer, cytokines can exhibit both protumor and antitumour effects. Some cytokines, such as IL-6 and TNF-alpha, promote tumor growth by stimulating cell proliferation, inhibiting cell death, and promoting angiogenesis (Cendrowicz et al., 2021). Conversely, other cytokines, such as IFN-gamma and tumor necrosis factor-beta (TNF-beta), exhibit antitumour effects by promoting cell death and activating immune cells to attack cancer cells. Macrophages play a key role in the early stages of cancer and help recognize and eliminate cancer cells. However, as the tumor grows, macrophages can become “reprogrammed” to adopt a protumor phenotype. These “tumor-associated macrophages” (TAMs) promote tumor growth by secreting cytokines and growth factors that stimulate cell proliferation and angiogenesis, suppress the immune response, and remodel the extracellular matrix to promote metastasis (Shen et al., 2018). Targeting cytokines and macrophages is a promising strategy for cancer therapy. For instance, drugs that block cytokine signaling, such as anti-IL-6 or anti-TNF-alpha antibodies, are currently undergoing clinical trials for various types of cancer. Additionally, there is growing interest in developing therapies that target TAMs, either by depleting them or reprogramming them to adopt an antitumour phenotype (Wei et al., 2023).
4. Current challenges:
Investigating the complex interplay between cytokines and macrophages in inflammation poses several significant challenges. First, the sheer diversity of cytokines involved complicates the identification of specific actors in individual cases. Overlapping and sometimes redundant functions of certain cytokines further enhance this complexity, demanding sophisticated tools for discerning their unique contributions (Cortés et al., 2023).
Determination of the concentrations of many cytokines in biological samples necessitates highly sensitive assays, such as flow cytometry or ELISA, for accurate quantification. Macrophages, a heterogeneous cell population, present another critical hurdle. The diverse tissue-specific phenotypes of these viruses exhibit distinct gene expression profiles and functional properties, necessitating tailored isolation techniques. Fluorescence-activated cell sorting (FACS) or magnetic-activated cell sorting (MACS) have become essential tools for differentiating these nuanced subsets (S. Chen et al., 2023). Maintaining macrophage viability throughout an experiment, especially during cytokine stimulation studies, can be delicate due to the high sensitivity of macrophages to environmental changes and the tendency for rapid activation or apoptosis. Deciphering the intricate signaling pathways activated by cytokines within macrophages is yet another challenge. Western blotting and gene expression profiling have become crucial tools for revealing the molecular cascades triggered by specific cytokines, allowing for a deeper understanding of their downstream effects (Oishi & Manabe, 2018). Both cytokines and macrophages exhibit pronounced heterogeneity, further adding layers of complexity to their roles in inflammation (Leone et al., 2023). Understanding the tissue-specific nuances of different macrophage subsets and the diverse effects of cytokine combinations is crucial for dissecting their precise contributions to the inflammatory process. Additionally, cytokine storms, characterized by a hyperactive immune response with excessive cytokine production, pose a potentially fatal threat, leading to widespread tissue damage and organ failure. Elucidating the mechanisms triggering such events holds immense clinical significance(Han et al., 2021). Finally, immunosenescence, the age-related decline in immune function, further complicates the picture. Understanding how these changes impact cytokine profiles and macrophage polarization patterns is vital for developing effective strategies to combat age-related infections, cancers, and autoimmune diseases. By addressing these significant challenges, researchers can elucidate the intricacies of cytokine–macrophage interactions in inflammation, paving the way for novel therapeutic interventions and improved human health outcomes (Kany et al., 2019b).
5. Future strategies:
The emergence of humanized animal models constructed by engrafting human cells or tissues into nonhuman hosts has revolutionized biomedical research and drug discovery (Chuprin et al., 2023). These models offer unparalleled advantages over traditional in vitro or in silico approaches by enabling the investigation of human pathophysiology in vivo, evaluating potential therapeutic efficacy, and providing a more accurate representation of disease progression than conventional methods. By serving as a preliminary testing ground for new drugs, humanized models can streamline the drug development process by identifying potential safety or efficacy concerns early on. Continued advancements in technology hold promise for the creation of even more sophisticated and precise humanized models, potentially paving the way for the development of more effective disease treatments (Ma et al., 2024).
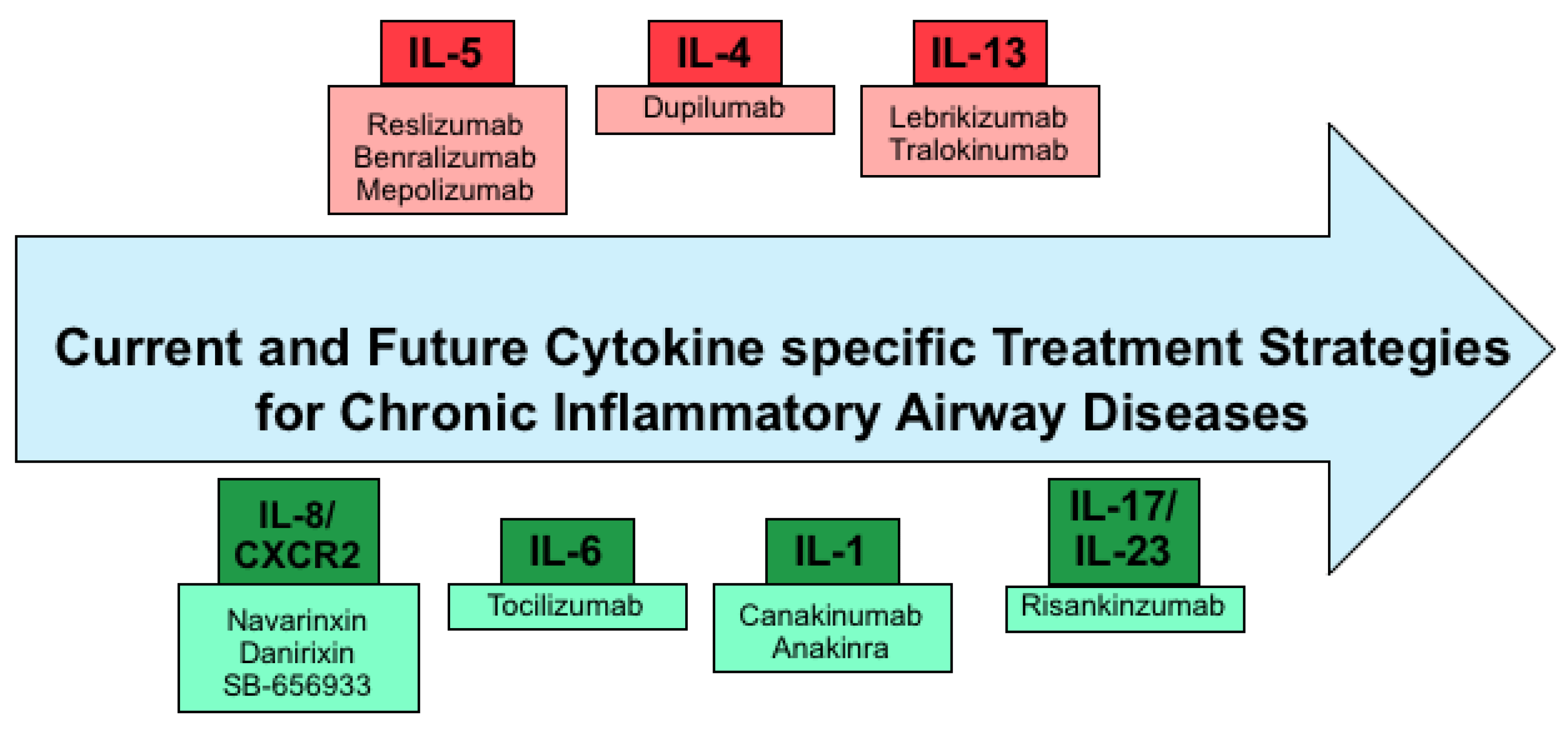
Figure 4: Current and future specific treatment strategies of cytokines. [source: (Garth et al., 2018)].
Single-cell sequencing, while a powerful tool for understanding cellular heterogeneity, faces several challenges. Isolation of individual cells from complex tissues or rare cell populations can be difficult, impacting subsequent analysis. The quality and quantity of isolated cells significantly influence the accuracy and reliability of the generated data. Additionally, the high-dimensional nature of single-cell sequencing data necessitates a meticulously designed analysis pipeline to manage technical variability and biological heterogeneity. Amplification, required to generate sufficient DNA or RNA for sequencing, introduces potential biases that can distort results, highlighting the importance of robust quality control measures. Macrophage polarization plays a pivotal role in regulating inflammation. Inducing M2 polarization through IL-4 and IL-13 has potential for mitigating inflammation in diseases such as asthma (Apeku et al., 2024). Emerging immunomodulatory therapies, such as senolytics and immune checkpoint inhibitors, offer promising avenues for combating immunosenescence. Senolytics selectively eliminate senescent cells, which contribute to age-associated inflammation, while immune checkpoint inhibitors enhance immune function by negating immunosuppressive signals. In the context of COVID-19, cytokine inhibitors such as tocilizumab and baricitinib have shown efficacy in managing cytokine storms by targeting specific proinflammatory cytokines. Despite the identification and study of numerous cytokines and macrophage subsets, the landscape has not been fully elucidated. Future research should focus on discovering novel entities and elucidating their roles in inflammation and the immune response (Pérez & Rius-Pérez, 2022). Dysregulated cytokine production is a hallmark of chronic inflammatory diseases. Deciphering the regulatory mechanisms governing cytokine production in macrophages and other immune cells provides fertile ground for identifying novel therapeutic targets. Similarly, the diverse spectrum of macrophage polarization states warrants further investigation to identify specific targets for modulating inflammation in various diseases. While the pivotal roles of cytokines and macrophages in inflammation and the immune response are well established, their specific contributions to many diseases remain unclear. Current imaging techniques provide limited information about cytokine and macrophage activity in vivo (Pérez & Rius-Pérez, 2022). The development of novel imaging modalities with the potential to visualize these activities in real time holds immense promise for advancing our understanding of inflammation and the immune response, paving the way for the development of more targeted and effective therapeutic strategies.
6. Conclusion:
In conclusion, macrophages exhibit remarkable heterogeneity and play a crucial role in both innate and adaptive immunity. The diverse activation states of M1 and M2 macrophages contribute to tissue repair, inflammation resolution, and disease progression depending on the specific context. Cytokines, signaling molecules produced by macrophages and other immune cells, further orchestrate these complex processes. Understanding the intricate interplay between cytokines and macrophages in inflammation holds immense potential for developing novel therapeutic interventions for various diseases, including cancer, autoimmune disorders, and chronic inflammatory conditions. Future research efforts should focus on deciphering the regulatory mechanisms governing cytokine production and macrophage polarization, identifying novel therapeutic targets, and developing advanced imaging modalities to visualize these activities in vivo. By addressing these challenges and revealing the secrets of cytokine–macrophage interplay, we can pave the way for a healthier future.
Funding: This research was unfunded by any public, commercial, or not-for-profit agency.
Ethical Approval and Consent to Participate: Not applicable.
Guarantor: The article’s full responsibility lies with PY, who is the corresponding author and the third author on the list.
Author contributions: SA, PS, SS, AK, and VR were responsible for manuscript conceptualization, writing - original draft, consent, and sample collection. PY participated in the writing, review and editing.
Acknowledgements: Due to space constraints, some pertinent publications could not be found.
Availability of data and materials: All the data have been truly cited in the articles.
Consent for publication: All the authors consented to the publication of this manuscript.

Open Access By Aditum Open Access Journals id licensed under Creative Commons Attribution 4.0 International License. Based On a Work at aditum.org